Механизмы и факторы энтоморезистентности древостоев
Уровень энтоморезистентности древостоев служит одним из ключевых элементов возможности возникновения вспышек массового размножения, а, следовательно, и механизма реализации вспышек, так как совершенно очевидно, что при высоком уровне энтоморезистентности возникновение вспышки массового размножения лесных насекомых-филлофагов невозможно, вследствие низкой выживаемости популяции.
Поэтому интерес к разработке этого направления исследований всегда был достаточно высок (Гримальский, 1971; Ильинский, 1959, 1960; Распопов, 1961; Руднев,1958, 1962; Воронцов, 1978; Кожанчиков, 1951; Положенцев, 1951; Ханисламов, 1958-1960, Ханисламов и др., 1958 a, b, 1962; Feeny, 1970), но в последнее время особенно возрос (Колтунов и др., 1992, 1998; Колтунов, 1993, 1996; Рафес, 1989; Рубцова, Рубцов, 1984; Bryantetall., 1993; Fagerstrometall., 1982; Haukioja, Niemela, 1978; Herms, Mattson, 1992; Houg, Pimentel, 1978; Houston, Valentine, 1985; Meyer, Montgomery, 1987; Rossiteretall., 1988; Schultz, Leckowitz, 1986).
В целом, известно, что параметры энтоморезистентности древостоев детерминируются широким кругом факторов различного уровня: структурными особенностями древостоев, возрастом древостоев, породным составом, почвенно-эдафическими факторами, температурой, интенсивностью и продолжительностью засух, суховеев, степенью ослабленности древостоя различными факторами, такими как антропогенное (техногенное) воздействие, рубки леса, порослевое возобновление, и многие другие (Гримальский, 1971; Воронцов, 1978; Ильинский, 1959; Руднев,1958; Рубцова, Рубцов, 1984; Houston, Valentine, 1985; Liebholdetall., 1995).
В наиболее значительной степени изучались физиолого-биохимические свойства листьев и хвои как кормового субстрата, так как было понятно, что именно эти параметры могут регулировать качество корма для насекомых и их выживаемость.
Значение рН древесных растений, как критерия устойчивости к повреждению насекомыми-вредителями и болезнями известно, давно. Еще П. А. Положенцев (1953) показал, что рН прикамбиального кольца у поражающихся насекомыми-вредителями деревьев выше, чем у здоровых. Но последующие исследования показали обратные результаты. рН у непоражаемых растений был ниже, чем у пораженных корневой губкой (Негруцкий, 1955). Но у пораженного дуба, по данным Ханисламова (1958) рН был выше, чем у контрольных. У лиственницы также по мере ухудшения состояния рН деревьев повышалась (Ханисламов и др., 1958).
Известно, также, что рН сока листьев увеличивается с начала распускания листвы до полного созревания, а затем, по мере старения листвы, уменьшается. Конкретная динамика рН зависит от вида кормового растения и внешних условий (Васфилов, 1997). Кроме того, заметная динамика рН характерна даже для листьев березы в течение суток (Васфилов, 1997).
Динамика рН листьев березы была особенно широка в начале органогенеза листьев и заметно сужается к середине (Пономарев, 1997, 2004). Максимального значения рН листьев березы достигает поздно вечером, или рано утром, минимального – в полдень. Когда гусеницы непарного шелкопряда достигают старшего возраста, у березы минимальный уровень рН в полночь (Пономарев, 1997, 2004).
Таким образом, возможно, что питание гусениц непарного шелкопряда младшего возраста утром и вечером энергетически выгоднее из-за низкого показателя рН кишечника (Пономарев, 1997, 2004). Исследования показали, что именно предпочитаемые гусеницами участки листа (основание листа, середина, возле центральной жилки) (80% гусениц питаются ими) наименее кислые (Пономарев, 1997, 2004).
В условиях хорошего увлажнения дефолиированные непарным шелкопрядом древостои имели более низкую фотосинтетическую норму. В условиях водного дефицита (засухи) дефолиация увеличивала фотосинтетическую норму.
Скорость роста непарного шелкопряда 4 возраста увеличивалась после воздействия фактора абиотического стресса (засухи) на древостои и уменьшалась в ответ на дефолиацию крон.
Многими авторами было показано, что гусеницы лучше развивались при питании листьями и хвоей древостоев, ослабленных воздействием различных факторов. При вспышках массового размножения непарного шелкопряда первичные очаги возникали в ослабленных различными факторами древостоях (Васильева, 1972; Васильева, Плешанов, 1972, 1975; Галкин, 1975; Гримальский, 1959; Ивлиев, 1960, 1966; Рожков, 1965, 1981; Руднев, 1958, 1962; Ханисламов и др, 1959, 1960, 1962).
По мнению Д. Ф. Руднева, (1958, 1962) изменение качества пищи под влиянием почвенных и антропогенных факторов является одной из главных причин колебаний численности насекомых-фитофагов. По данным В.И. Гримальского эфирные масла физиологически здоровых сосен для гусениц хвоегрызущих насекомых являются токсичными, но в условиях засухи и дефицита азота в почве интенсивность смоловыделения значительно уменьшается (Гримальский, 1959). Соответственно, значительно возрастает и выживаемость гусениц хвоегрызущих насекомых при питании этой хвоей.
Но при этом совершенно очевидно, что, учитывая значительные площади очагов массового размножения непарного шелкопряда, все леса не могут быть постоянно ослабленными. Таким условиям удовлетворяют лишь отдельные, локальные участки леса вблизи населенных пунктов, или участки, расстроенные постоянным выпасом скота, другими антропогенными факторами, а, также, некоторые лесокультуры.
Как показали полученные нами результаты многолетних исследований, для успешной реализации вспышки массового размножения непарного шелкопряда вполне достаточно не постоянного ослабления устойчивости лесов, а временного (Колтунов и др., 1992; 1998;2010; Колтунов, 1993, 1996,2006). Роль этого фактора временного ослабления энтоморезистентности очень эффективно выполняет фактор абиотического стресса (весенне-летние засухи).
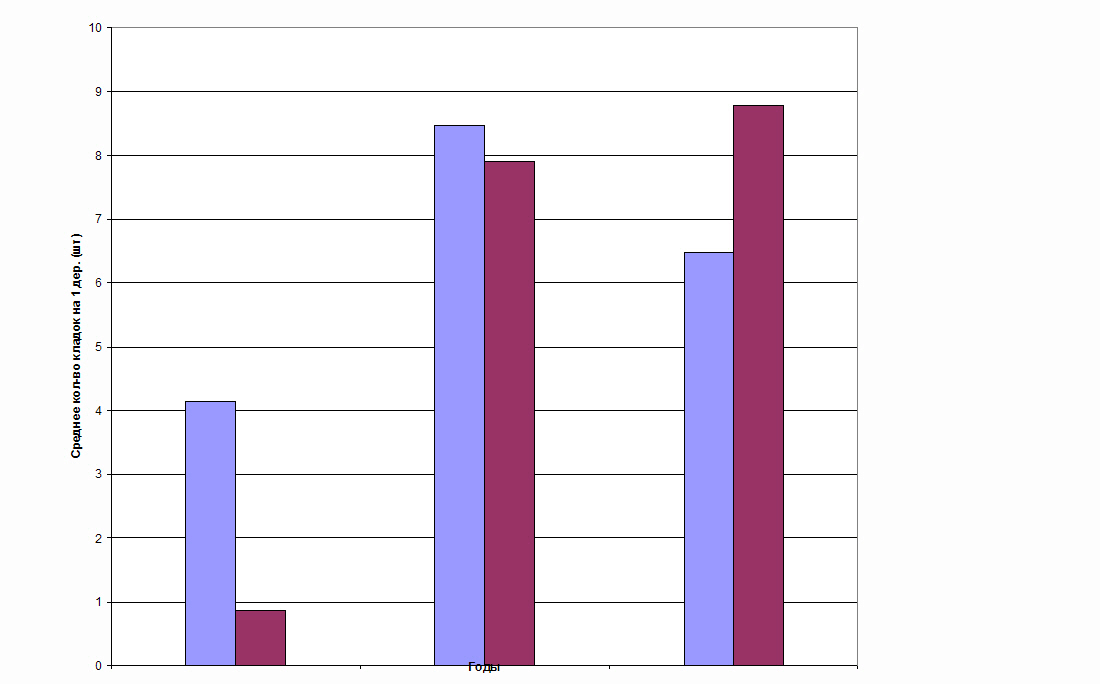
Рис.1. Динамика заселенности березы кладками непарного
шелкопряда в период вспышки массового размножения
в течение 3 лет в насаждении с низкой (слева) и высокой
(справа) энтоморезистентностью. Челябинская обл.
Как показали результаты исследований, в березовых лесах с высокой и низкой энтоморезистентностью в течение вспышки массового размножения нет заметных отличий как по абсолютным показателям заселенности, так и по характеру динамики (рис.1). Единственным отличием было то, что в низкорезистентном березняке в первый год вспышки абсолютная заселенность кладками всегда выше, чем в высокорезистентных древостоях. В остальные годы вспышки показатели абсолютной заселенности древостоев кладками непарного шелкопряда совершенно не отличаются.
Следовательно, по показателям плотности заселения березняков кладками непарного шелкопряда, или характеру динамики их заселенности совершенно невозможно отличить насаждения с низкой (дефолиация до 100 %) и высокой (дефолиация до 20-30 %) энтоморезистентностью. Это вполне объяснимо, так как критерии предпочтения древостоев самками имаго совершенно иные (Колтунов, 1993, 1996;2006; Колтунов и др., 1998,2010). Они совершенно не совпадают с критериями предпочтения древостоев гусеницами этого фитофага.
Это свидетельствует, также, о том, что в очагах сильная дефолиация крон всегда осуществляется собственной микропопуляцией непарного шелкопряда, а не мигрантами, хотя, как показано нами ранее, уровень миграционной активности гусениц в период вспышки очень высок (Колтунов и др., 1998).
Это особенно очевидно для вспышек массового размножения сибирского шелкопряда в темнохвойных лесах Ханты-мансийского округа, часть которых совершенно не затронута антропогенным воздействием (в том числе и рубками леса), но, тем не менее, подверглась дефолиации, наряду с участками, где проводилась рубка леса (Колтунов, 1996; Колтунов и др., 1996).
При этом важную роль во временном ослаблении лесов выполняют (как показано нами в главе 5) не только полные весенне-летние засухи (май-август), но и частичные (только весенние, которые чаще встречаются, чем полные засухи). Особенно важное значение для реализации вспышки имеют майские засухи, когда гусеницы младшего возраста особенно чувствительны к воздействию неблагоприятных факторов окружающей среды. Роль летних засух меньше, особенно – позднелетних (август) (Колтунов, 2006).
Поэтому совершенно очевидно, что лесные фитоценозы, в которых происходит возникновение периодических вспышек массового размножения насекомых-филлофагов, не являются постоянно ослабленными. Они могут возникать и в труднодоступных участках леса, где исключено любое воздействие антропогенных факторов. Наиболее часто постоянно ослабленными является лишь небольшая часть древостоев, в которых нередко и происходит образование первичных очагов, формирование стаций переживания (Воронцов, 1978; Знаменский, 1987; Ханисламов и др., 1962; Распопов, 1961, 1970, 1973). Но, в основном, лесные фитоценозы являются лишь временно, обратимо ослабленными в результате воздействия фактора абиотического стресса (весенне-летних засух).
Нами показано, что в условиях лесостепи Зауралья в период отдельных вспышек массового размножения непарного шелкопряда воздействие фактора абиотического стресса иногда наблюдается в течение 7 лет, ежегодно. После же восстановления нормальных климатических условий их энтоморезистентность быстро восстанавливается.
Эта гипотеза, по нашему мнению, достаточно адекватно решает проблему того, каким образом могут возникать вспышки массового размножения в лесах, совершенно недоступных любому виду хозяйственного воздействия (леса в труднодоступных районах Красноярского края), которые явно не могут быть отнесены к постоянно ослабленным лесам (Баранчиков и др., 2001).
Тогда становится понятно и то, каким образом происходит реализация вспышки массового размножения непарного шелкопряда в ареале дуба, когда нет синхронизации эруптивной фазы с весенне-летней засухой. В этих условиях подъем численности синхронизирован с засухами, а эруптивная фаза может реализоваться и на фоне нормальной влажности, за счет инерционных параметров популяции с возросшим вспышечным потенциалом. Тем не менее, внимательный анализ показывает, что все-таки, в большинстве случаев, многие вспышки массового размножения непарного шелкопряда в Южной Европе в дубовых лесах хорошо синхронизированы с засухами. Именно в случае хорошей синхронизации площади очагов особенно значительны.
Но ключевым популяционным фактором в системе взаимоотношений: «дерево-насекомые», позволяющим успешно реализовать вспышку массового размножения, по нашему мнению, является не плодовитость, и ее рост по мере возрастания численности популяции (на что обращают особенное внимание некоторые авторы (Васильева, 1972), а только баланс между выживаемостью и смертностью. Именно резкий рост выживаемости, а не плодовитости, обеспечивает быстрое возрастание численности и экологической плотности популяции непарного шелкопряда. И, соответственно, именно этот параметр и детерминирует возможность успешной реализации вспышки. Это подтверждается и мнением других авторов (Рожков, 1981).
Именно очень низкая выживаемость популяции приводит к тому, что, несмотря на высокую заселенность древостоев кладками и хорошую отрождаемость, до старших возрастов доживает очень незначительное количество гусениц. В результате дефолиация крон остается очень незначительной (Колтунов, 1993, 1996; Колтунов и др., 1998).
В дальнейшем, по мере развития очагов, основным препятствием для роста площади очагов является фактор индуцированной резистентности древостоев. Нами показано, что в березовых лесах Зауралья сильной повторной дефолиации крон, практически, не встречается (Колтунов и др., 1992; Колтунов, 1993, 1996; Колтунов и др., 1998). Но это не является отличительным признаком этого вида (березы), так как в условиях возникновения очагов массового размножения насекомых-филлофагов летне-осеннего экологического комплекса, повторная сильная дефолиация наблюдается (Соколов, 2005). Хотя площадь таких повторно дефолиируемых березняков всегда незначительна и не превышала (в период последней вспышки массового размножения насекомых-филлофагов летне-осенней экологической группы 5% от общей площади очагов.
Известно, что механизм индуцированной энтоморезистентности обычно функционирует при повреждении крон весенне-летними видами насекомых-филлофагов и не функционирует при повреждении крон летне-осенними видами насекомых-филлофагов (Мамаев, Рубцов, Уткина, 2001, 2002; Иерусалимов, 2004).
Но в системе: «дуб-непарный шелкопряд» определенная часть древостоев подвергается повторной сильной дефолиации (Воронцов, 1978; Рубцов, Рубцова, 1984; Старцев, 1983; Gottschalk, 1993 ).
Таким образом, вероятно, эффективность функционирования механизма индуцированной резистентности тесно связана с видовыми особенностями древесных растений. Хотя следует отметить важную деталь: площадь повторно дефолиируемых дубняков обычно, по данным различных авторов, не очень велика (Воронцов, 1978; Рубцов, Рубцова, 1984; Старцев, 1983; Gottschalk, 1993).
Интересно, что, по данным ряда авторов, кормовая ценность листвы неравнозначна даже, в зависимости от высоты кроны деревьев. Известно, что, в первую очередь подвергается дефолиации верхняя часть кроны, как наиболее предпочитаемая многими видами насекомых-фитофагов (Воронцов, 1978; Кононова, 1964). Так, Кононовой (1964) показано, что при питании гусениц непарного шелкопряда листьями дуба из нижней части кроны выживаемость была ниже (6%), чем верхней (13%). Но самой высокой была выживаемость гусениц при питании их молодыми листьями после заморозков (15%).
Таким образом, анатомо-морфологические особенности листьев оказывают влияние на насекомых и качество пищи. Кроме того, воздействие заморозков вполне уместно рассматривать как воздействие фактора абиотического стресса, который заметно изменяет кормовую ценность листьев в сторону улучшения.
Известны, также, данные о том, что полив деревьев и внесение минеральных удобрений снижают выживаемость непарного шелкопряда (до 26% при 42 % в контроле) (Кононова, 1964). Гримальский (1971) позже подтвердил, что повышение смертности гусениц в холодную, дождливую погоду обусловлено повышением кислотности хвои. На старопахотных почвах она была выше (17,07; 19,94 мг/экв) по сравнению с лесными почвами (11,81; 15,92 мг/кв.). Гибель гусениц при питании хвоей сосны, растущей на старопахотных почвах, также была выше. Одновременно, как показали результаты, хвоя была обеднена питательными веществами: сахарами и азотом.
М. Д. Тарануха (1952) указывал, что древостои с повышенным содержанием растворимых углеводов (сахаров) и соответствующим количеством азотистых соединений в листьях способствуют развитию и росту плодовитости непарного шелкопряда.
Но, все-таки, устойчивость сосны к повреждению хвоегрызущими насекомыми-филлофагами определяется не столько содержанием в хвое питательных веществ, как показано В.И. Гримальским (1971). Мы предполагаем, что различия в кормовой ценности одного вида деревьев в разных условиях местопроизрастания могут объясняются не разным содержанием в них питательных веществ, а разным содержанием веществ, препятствующих питанию, обладающих антифидантными свойствами.
Но, согласно экспериментальным данным, содержание дубильных веществ в сосняках на старопахотных почвах 2,54%, тогда как в сосняках на лесных почвах: 1,88%. Следовательно, они не оказывают влияния на выживаемость гусениц непарного шелкопряда, так как все очаги возникали в сосняках на старопахотных почвах (Гримальский, 1971).
По содержанию эфирных масел в хвое авторами были получены следующие данные: в хвое сосны на старопахотных почвах – 0,10 мл/100 г хвои, в хвое сосны, растущей на лесных почвах – 0,42 мл/100 г хвои. Но смертность гусениц, питающихся хвоей сосны, растущей на лесных почвах, была значительно выше, чем на старопахотной.
Как показали результаты, с увеличением содержания эфирных масел в хвое, возрастала также смертность гусениц соснового шелкопряда и сосновой совки (Гримальский, 1971). Но, по данным анализа, с возрастом гусениц возрастала и их устойчивость к эфирным маслам хвои.
Искусственная обработка хвои эфирными маслами в лабораторных условиях приводила к массовой гибели гусениц, питающихся ею (Гримальский, 1971).
Дальнейшее развитие исследований автора в этом направлении позволили сформировать автору новую гипотезу: для устойчивости против хвоегрызущих вредителей основное значение имеет, собственно, не абсолютное значение содержания эфирных масел в хвое, как таковое, а интенсивность их выделения после повреждения гусеницами. А интенсивность смоловыделения зависит не столько от содержания эфирных масел, сколько от тургора клеток хвои. Он же, в свою очередь, зависит от содержания воды в клетках. Понятно, что хвоя сосны на старопахотных почвах имела пониженную влажность. Поэтому и смоловыделение было слабым (Гримальский, 1971).
Интересно, что интенсивность смоловыделения резко снижается уже через 15-30 мин после срезания веток. Вот почему многие данные современных лабораторных экспериментов по выкармливанию гусениц на срезанных ветках и на деревьях совершенно не совпадают.
Позже исследованиями D’Otto, W. Hackbarth (1967) было показано, что интенсивность смоловыделения у хвойных растений на бедных почвах падает.
При этом многие авторы отмечали роль вторичных соединений как химической основы резистентности кормового субстрата. Наэтурольрядавтороввыдвинулфенолы (Rossiter et all., 1988; Herms, Mattson, 1992; Bryant et all., 1993; Kinney et all., 1997). Кроме того, было установлено, что танины связывают белки, снижая таким образом, перевариваемость белков в организме насекомых и снижая их пригодность для насекомых (Sunnerheimetall., 1988). Было обнаружено, что физиологическая зрелость листьев через год после дефолиации крон деревьев была ниже, чем защищенных от дефолиации.
Таким образом, в целом, ответ дерева на фактор дефолиации складывается из ответов, вызванных непосредственно эффектом повреждения, изменений, следующих по иерархии и изменений, следующих из физиолого-биохимических изменений листьев.
Многие исследования показали, что в листве кормовых растений после размножения насекомых-фитофагов повышается количество фенолов и других веществ с антибиотическими свойствами (Рафес, 1981; Haukiojaetall., 1978, 1985; Haukioja, 1990; Fagerstrometall., 1982, Колтунов, Хамидуллина, 2012, 2013).
По мнению П.М. Рафеса, такое ухудшение кормовых свойств растений, вероятно, происходит в процессе их репарации за счет компенсаторного фотосинтеза (Рафес, 1989). В результате этого часть гусениц погибает, происходит, также, задержка окукливания.
Но, в дальнейшем, эта гипотеза пока не получила однозначного подтверждения. Учитывая очень эффективное функционирование механизма индуцированной энтоморезистентности (после 30% и более дефолиации) у березы (Колтунов,1993, 1996, 2006; Колтунов и др., 1992, 1998, 2010), мы предполагаем, что механизм такой защиты, безусловно, существует, но он не обязательно может быть связан с фенолами. Многие другие вещества могут выполнять защитную функцию. Это направление исследований нуждается в дальнейшем развитии.
Было показано, что устойчивость древостоев к сосновой пяденице не коррелирует с величиной среднего радиального прироста древостоев (Суховольский, Артемьева, 1992). Позже эти результаты были подтверждены С.В. Соколовым в системе: «береза – летне-осенний экологический комплекс насекомых-фитофагов» (Соколов, 2005).
Установлено, что чем меньше высокочастотная составляющая радиального прироста, тем сильнее повреждалось дерево (Суховольский, Артемьева, 1992). В неповрежденных древостоях высокочастотные характеристики выше. Авторы не объясняют, почему с увеличением частоты ВЧ-колебаний прироста устойчивость древостоя к вредителям растет.
Позже, такую же работу по анализу взаимосвязи энтомотолерантности березы и ВЧ-составляющей прироста провел С. В. Соколов и подтвердил известные ранее данные о взаимосвязи этих параметров (Соколов, 2005).
Установлено, что общее количество азота, сахаров, белков, крахмала, лигнина и гемицеллюлозы было ниже, а концентрация фенолов была выше в листьях, дефолиированных в предыдущем году, по сравнению с листьями, специально защищенными от дефолиации (Haukiojaetall., 1985; Rossiteretall., 1988).
Масса куколок положительно коррелировала с концентрацией азота и сахара в листьях.
Таким образом, очевидно, что дефолиация листьев в предыдущем году изменяет биохимический состав листьев в следующем году, что отражается на его трофических свойствах. Авторы предполагают, что это обусловлено задержкой биохимического созревания листьев.
В целом растения имеют много разнообразных механизмов защиты от насекомых-вредителей. Среди них одним из самых важных являются индуцированные повреждением насекомых-фитофагов биохимические изменения состава листьев, которые становятся менее приемлемыми для вредителей. Это может происходить либо за счет увеличения токсичности субстрата, либо за счет снижения его кормовой ценности (Haukioja,1990; Herms, Mattson, 1992; Kaitaniemietall., 1998).
Многократная дефолиация древостоев непарным шелкопрядом, как правило, отсутствует. Как показали полученные нами результаты, это происходит вследствие эффективного функционирования у березы механизма индуцированной энтоморезистентности (Колтунов, 1993, 1996, 2006; Колтунов и др., 1992, 1998, 2010). Несмотря на высокую численность яйцекладок непарного шелкопряда в древостоях, полностью, или сильно дефолиированных в предыдущем году, на следующий год повторной сильной дефолиации не обнаруживается. Она не превышает 15-20%. (рис. 2). Это происходит вследствие резкого
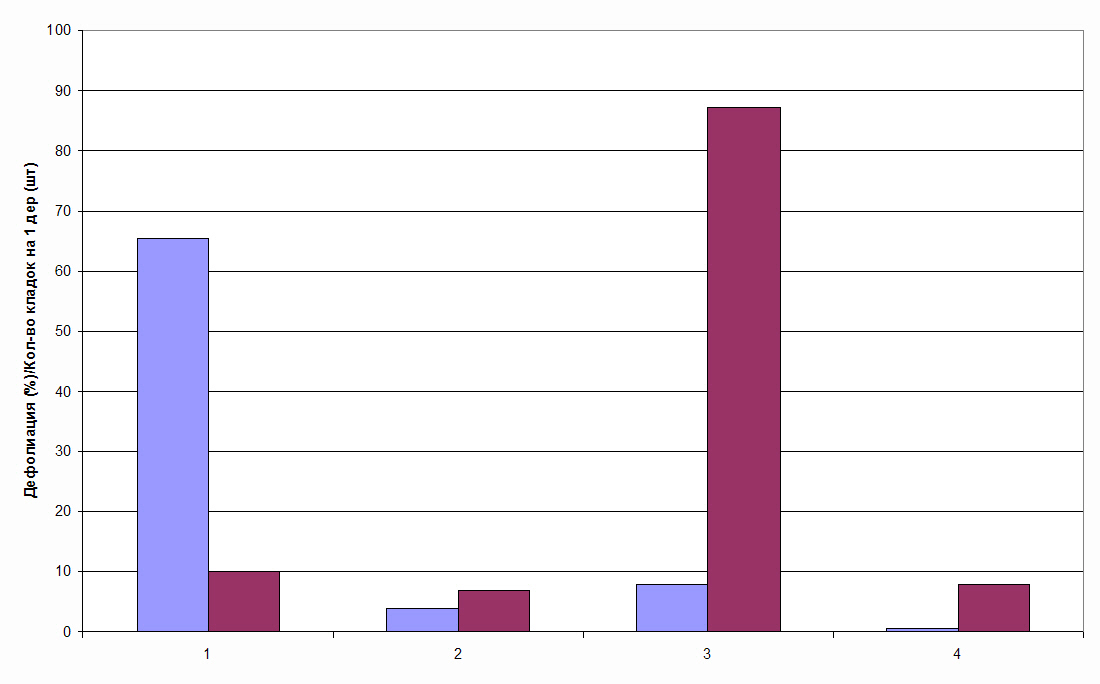
Рис.2. Влияние фактора индуцированной энтоморезистентности на повторную дефолиацию крон березы в очаге массового размножения непарного шелкопряда в Челябинской обл. Слева 1988 г., справа – 1989 г. 1,3 –дефолиация; 2,4 – кол-во кладок на 1 дерево (1-2; 3-4). 1 – фактор индуцированной энтоморезистентности присутствует; 3 – отсутствует.
снижения выживаемости гусениц при питании листьями ранее дефолиированных деревьев. Как видно из рис. 4.1 при дефолиации менее, чем 20-30% механизм индуцированной энтоморезистентности не инициируется, так как на следующий год наблюдается очень сильная дефолиация крон. Как показано нами ранее (Колтунов, 1993, 1996, 2006; Колтунов и др., 1992; 1998, 2010), а , также другими авторами, этот механизм эффективно функционирует в течение 3-4 лет.
Исследования показали, что гусеницы, дающие самок, были более чувствительны к качеству кормового субстрата.
При этом только питание гусениц младших возрастов молодой хвоей темнохвойных может идти нормально.
В ответ на содержание в пище аллелохимиков у непарного шелкопряда была обнаружена индукция синтеза трипсина, В-глюкозидазы (Lindroth, 1988), и глютатион- S-трансферазы (Periс-Matarugaetall., 1997).
Быстрое снижение содержания азотистых веществ в листьях происходит в годы засухи, а повышенное содержание азота в пище, или его колебание приводит к снижению его ассимиляции гусеницами (Stocknoff, 1993).
P. Feeny (1970) предполагает, что выживаемость листогрызущих насекомых связана с возрастом, обводненностью и содержанием танинов в листьях.
Haukioja, Niemela (1978) предположили, что основной механизм защиты против дефолиации – содержание фенолов в тканях растений. В дальнейшем это, к сожалению, не нашло однозначного подтверждения в экспериментах и в 1989 г. авторы отказались от этой гипотезы. При этом они высказали другое предположение, что, возможно, более важно для этой функции содержание углеводов в листьях.
Н. М. Эдельман (1963) показала, что требовательность гусениц непарного шелкопряда к биохимическому составу корма в младших и старших возрастах различается. Совпадение сезонных изменений биохимического состава с требованиями насекомых к пище на всех этапах онтогенеза является одной из причин вспышек массового размножения.
Известно также, что при питании листвой в результате разрушения вакуолей происходит образование белково-танниновых комплексов. Предполагается, что в этом и заключается защитная роль танинов от дефолиации (Schultz, Leckowitz, 1986). Но у непарного шелкопряда предпочитаемыми породами являются именно те, которые содержат большое количество танинов . Это: Salicaceae, Betulaceae. И именно при высоком значении рН корма, содержащего танины, гусеницы достигали большего веса при меньшем потреблении пищи (Govenor, etall, 1997). Как известно, белково-таниновые комплексы образуются в кислой среде. При рН>9,0 они распадаются (Schultz, Leckowitz, 1986).
Широко известно, что рН кишечника гусениц имеет высокое значение (Кузнецов, 1948; Колтунов, Фадеев, 1981). Очень важно и то, что в процессе биохимической коэволюции и протеазы кишечника гусениц также имеют максимальную ферментативную активность в области того значения рН, которое имеет рН кишечника хозяина (Колтунов, Фадеев, 1981).
Многие исследователи пришли к выводу о том, что наиболее часто крупные вспышки массового размножения насекомых-филлофагов возникают в определенных типах лесных фитоценозов (Воронцов, 1978; Ильинский, 1959; Распопов, 1961; Ханисламов, 1958; Mattson, 1997 ). Они имеют ряд характерных особенностей: древесный ярус состоит из одного, или нескольких видов древесных растений, все из которых являются чувствительными при повреждении вспышечными видами насекомых (Mattsonetall., 1991; Mattson, 1997). Это часто леса, растущие на бедных, недостаточно обеспеченных водой почвах. Древесный ярус часто находится в угнетенном состоянии.
Наиболее хорошим примером таких взаимоотношений являются системы: «темнохвойные леса – сибирский шелкопряд», «дуб-непарный шелкопряд». В многовидовых лесных сообществах вспышки массовых размножений насекомых-филлофагов редки и являются нормальной частью динамики сукцессионных процессов и не наносят эколого-экономического ущерба лесным фитоценозам.
Mattson (1995, 1997) считает, что такие крупные вспышки массового размножения происходят только в условиях, где факторы внешней среды ограничивают длительную жизнеспособность древесных растений. Например, таким фактором могут быть засухи, которые ослабляют древесные растения. Известно, также, что в дубравах лесостепи таким фактором часто являются, также, особенно морозные зимы, ослабляющие дуб (Рубцова, Рубцов, 1984).
В таких сугубо маргинальных условиях роста древесные растения постоянно, или часто подвергаются стрессовому воздействию засух, недостатку минерального питания. Имеется, также, много различных антропогенных факторов, на которые остро реагируют лесные фитоценозы. Это – сильная рекреационная и пастбищная дигрессия лесов (Колтунов, 1993, 1996, 2006; Колтунов и др., 1998, 2010) техногенное загрязнение, дефрагментация лесов, деградация подроста и подлеска и травяного яруса вследствие интенсивной антропогенной дигрессии. Это быстрое возрастание уровня СО в атмосфере, которое может быть очень значимым фактором, изменяющим фундаментальный характер как растительной ткани, так и путей взаимодействия с другими компонентами биогеоценоза, контролирующими ключевые изменения структуры лесных сообществ. Долговременные последствия таких изменений зависят от индивидуальных особенностей реакции древостоев и насекомых-фитофагов на увеличение содержания СО2 в атмосфере (Ayres, 1993; Vitousek, 1994).
Следует, также, учесть, что, кроме прямого эффекта рост содержания СО2 в атмосфере, вместе с ростом уровня других газов, продуцируемых зелеными растениями, может привести к повышению глобальной температуры планеты. Это же, в свою очередь, может инициировать локальные изменения климата и путей взаимодействия растений с фитофагами.
Учитывая происходящие глобальные изменения мирового цикла азота, и, как следствие эвтрофикацию экосистем (Vitousek, 1994), следует иметь в виду, что такое огромное увеличение азота может внести значительные изменения в экосистемы, вследствие изменения роста растений. Это, в свою очередь, может привести к значительным изменениям условий обитания насекомых-фитофагов и взаимоотношениям в системе: «дерево-насекомые».
Увеличение уровня токсичных поллютантов, а, также, озона в атмосфере может иметь самые серьезные глобальные последствия для лесных биогеоценозов, так как озон губительно действует на фотосинтез растений, повреждая хлоропласты. Это может повысить чувствительность растений к насекомым-филлофагам (Riemer, Whittaker, 1989).
Исследования показали, что древесный ярус состоит из чувствительных к дефолиации деревьев и резистентных (в разной степени). Одновидовые сообщества деревьев обычно более чувствительны к дефолиации, чем смешанные древостои (Bessetall., 1961; Gottschalk, Twery, 1995; Valentine, 1981).
Дифференциация древостоев на резистентные и чувствительные привела ряд исследователей к идее организации лесных культур в зоне обитания и вспышек массового размножения непарного шелкопряда только из высоко резистентных видов, за счет удаления чувствительных к повреждению непарным шелкопрядом видов (Clementetall., 1936; Rostian, Ruggles, 1941).
При этом, нужно отметить очень важный аспект. Фактически, обсуждая уровень энтоморезистентности, или чувствительности той, или иной породы деревьев, на самом деле, имеется в виду лишь резистентность, или чувствительность древостоя только по отношению к повреждению непарным шелкопрядом, т.е. в очень узком смысле. Совершенно очевидно, что древостой может быть высоко резистентным к одному виду насекомых и чувствительным к другому виду. Это, к сожалению, может быть одним из весомых аргументов против реальности технологии защиты лесов США от непарного шелкопряда, рассматриваемой рядом авторов.
Кроме того, мы не можем согласиться и с самой идеей в целом, так как она потребует массовой вырубки ценнейших дубовых лесов, сосновых лесов, которые также повреждаются непарным шелкопрядом.
Также нельзя не иметь в виду огромные экономические затраты на вырубку лесов и лесопосадки нечувствительных к непарному шелкопряду видов. Кроме того, следует отметить, что точно такой же эффект, но только в более замедленном варианте, производит и непарный шелкопряд, повреждая только древостои, чувствительные к нему.
Таким образом, предлагаемая идея не имеет реальных научных оснований для ее реализации. Значительно меньших экономических затрат потребует проведение лесоэнтомологического мониторинга в местах обитания непарного шелкопряда по степени устойчивости лесов к повреждению непарным шелкопрядом, картирование зон с высокой чувствительностью и проведение активных мероприятий по защите леса при прогнозировании сильной дефолиации.
Многие авторы к чувствительным к непарному шелкопрядом лесам относят леса, расположенные в сухих местообитаниях, в степной и лесостепной зонах (Распопов, 1961; Амирханова, 1963), где древостои состоят, в основном, из медленно растущих, чувствительных видов и часто подвергаются воздействию природных и антропогенных неблагоприятных факторов (Houston, Valentine, 1985).
Соответственно, эти же авторы (Houston, Valentine, 1985) описывают резистентные к этому фитофагу древостои как древостои, растущие на склонах, в поймах, где влажность не является лимитирующим фактором, состоящие из большого количества разных видов.
Отметим, что ранговая структура факторов, детерминирующих чувствительность или резистентность древостоев к дефолиации в разных лесорастительных условиях заметно различается. Так, в засушливых условиях степи и лесостепи дефицит влаги является основным фактором, детерминирующим энтоморезистентность древостоя к дефолиации непарным шелкопрядом (Колтунов, 1993, 1996, 2006; Колтунов и др., 1992, 1998, 2010) в березовых колковых лесах. Заметим, что, по логике авторов, цитируемых выше, единственным признаком чувствительности, или резистентности к непарному шелкопряду является порода дерева, что совершенно недостаточно для понимания механизмов энтоморезистентности.
Как показали полученные нами результаты, даже в одновидовых сообществах, состоящих только из чувствительного к дефолиации вида (березы) вся популяция дифференцируется на древостои с высокой чувствительностью, средней, слабой и резистентные (Колтунов, 1993, 1996, 1998). Здесь следует еще раз повторить, что, несмотря на то, что береза относится к высокочувствительным к дефолиации непарным шелкопрядом видом, часть древостоев в ценопопуляции обладают в период вспышки высокой резистентностью. Механизм их энтоморезистентности подробно проанализирован нами в главе . Кратко он сводится к тому, что, вследствие определенных лесорастительных условий эти древостои очень слабо реагируют на фактор абиотического стресса (весенне-летние засухи), в отличие от группы чувствительных древостоев, которые сильно реагируют на этот абиотический фактор и быстро дефолиируются непарным шелкопрядом. Основной причиной резистентности, как уже указано выше, является быстрое изменение биохимического состава листьев у чувствительных древостоев, отреагировавших на стресс (засуху) и отсутствие таковых в группе временно резистентных древостоев.
Но при определенных условиях, позже и часть этих, ранее резистентных древостоев может стать чувствительной к дефолиации, если через определенный период времени влажность почвы в месте их произрастания заметно снизится (Колтунов, 1993,1996; Колтунов и др., 1998). Другая же часть, растущая в почвенных условиях, хорошо обеспеченных влагой, остается резистентной весь период вспышки массового размножения.
Таким образом, даже в одновидовых сообществах наблюдается значительная дифференциация по уровню энтоморезистентности древостоев, в зависимости от лесорастительных условий (почвенно-эдафических факторов). Здесь не обсуждается еще и проблема изменения энтоморезистентности, в зависимости от возраста, плотности древостоя и других факторов, что хорошо известно из многочисленных публикаций (Ильинский, 1958; Распопов, 1961; Воронцов, 1978). Не рассматриваем мы, также, и индуцированную энтоморезистентность, инициированную слабой дефолиацией в предыдущий год. А этот феномен очень перспективен для использования в защите леса, когда можно допустить слабую дефолиацию древостоев, которые затем, в течение всей вспышки будут иметь высокую энтоморезистентность к дефолиации непарным шелкопрядом. Но это можно использовать лишь относительно тех пород деревьев, которые не повреждаются непарным шелкопрядом вторично, со значительным уровнем дефолиации крон. Но, исходя из анализа публикаций, при нормальных лесорастительных условиях такими является большинство пород деревьев.
Заслуживает внимания и проблема влияния на уровень энтоморезистентности различных абиотических факторов, к числу важнейших из которых являются климатические условия. Очевидно, что засушливые условия заметно повышают чувствительность древостоев к дефолиации.
Учитывая все это, проблема энтоморезистентности более сложна и не должна рассматриваться лишь на уровне видов древостоев.
Известно, что выкармливание гусениц непарного шелкопряда листвой разного возраста оказывает значительное влияние на смертность, плодовитость и скорость онтогенеза (Кожанчиков, 1951; Houg, Pimentel, 1978; Meyer, Montgomery, 1987). Так, питание майской листвой обеспечивало более высокую скорость онтогенеза гусениц непарного шелкопряда, по сравнению с июньской листвой и прирост массы тела в течение 3 возраста (Андреева, 2000, 2002 a, b).
Как показали исследования, важнейшее значение для эффективности питания гусениц непарного шелкопряда имеет не только и не столько химический состав кормового субстрата, как таковой, но и рН этого субстрата (Андреева, 2002; Пономарев, 2004
). Так, показатель эффективности использования пищи был всегда выше при питании листвой с более высоким рН (Андреева, 2002). Известно, что рН клеточного сока тесно связана с интенсивностью фотосинтеза (Васфилов, 1997).
Практически всегда в насаждениях, дважды подвергшихся дефолиации насекомыми-филлофагами более, чем на 50%, происходит либо частичное, либо полное усыхание древостоя (Мамаев, Рубцов, Уткина, 2002; Иерусалимов, 2004).
Таким образом, проведенный нами обзор данных по проблеме энтоморезистентности показал, что, несмотря на длительный период и интенсивность исследований в этом направлении, имеющиеся результаты не приблизили понимание механизмов возникновения вспышек массового размножения. Это, по нашему мнению, однозначно свидетельствует о некоторых несомненных параметрах механизмов вспышек. Во-первых, очевидно, что механизм вспышек не связан с определенным количеством питательных веществ в листьях и хвое, хотя, в целом, и прослеживаются определенные закономерности. Поэтому дальнейшие поиски механизма в этом направлении крайне неэффективны. Во-вторых, параметры энтоморезистентности, совершенно, очевидно, детерминируются различными компонентами лесного фитоценоза разного рангового уровня, как показано нами ранее (Колтунов, 1993, 1996, 1998). Уже по этой причине невозможно выделить из сложной системы параметров один, или группу факторов одного ранга (например, содержание сахара, азота в листьях).
Поэтому исследование механизмов энтоморезистентности целесообразно продолжить на другом, системном уровне.
Перед тем, как изложить полученные нами результаты исследований механизмов энтоморезистентности, считаем целесообразным отметить основные, полученные нами ранее результаты, изложенные в предыдущих монографиях (Колтунов, 1993, 1998, 2006).
Было установлено, что даже в одновидовых сообществах (березовые леса) уровень энтоморезистентности березы колеблется в очень широком диапазоне. Основными параметрами варьирования энтоморезистентности были природные, а не антропогенные факторы. Вклад антропогенных факторов был значим при значительном уровне антропогенной трансформации лесных фитоценозов (с 4 стадии антропогенной трансформации, где основным фактором нарушенности была пастбищная дигрессия). Влажность почв также была важным фактором энтоморезистентности, так как в ксерофитных условиях уровень энтоморезистентности древостоев всегда был ниже, чем во влажных. С возрастом средняя степень дефолиации крон березы повышается. Уровень энтоморезистентности березы пушистой, в среднем, выше, чем у березы повислой, что может быть обусловлено и более влажными почвенными условиями, которые предпочитает береза пушистая.
Установлено, что, несмотря на длительную адаптацию к питанию березой у зауральской популяции, она, тем не менее, не является предпочитаемой породой у непарного шелкопряда, так как он оказывал явное предпочтение ольхе и осине и только после дефолиации их переходил на питание березой.
Одним из наиболее существенных результатов, полученных ранее, было то, что уровень энтоморезистентности березы в лесостепи Зауралья никак не был связан с обеспеченностью почв минеральными элементами. То есть, в отличие от результатов, полученных ранее Гримальским, предполагающим, что очаги возникают только в древостоях, растущих на бедных почвах (что, видимо, было характерно для лесокультур), вспышки массового размножения непарного шелкопряда возникали и на бедных и на богатых почвах. Но вначале вспышки очаги возникали только на бедных по минеральному составу почвах. Основной причиной, как показано нами ранее, было то, что древостои, растущие на относительно бедных почвах раньше откликались на воздействие фактора абиотического стресса (засуху), чем древостои, растущие на богатых почвах. Отклик на этот фактор стресса у древостоев, растущих на богатых почвах, был более замедленным, и наступал позже . После этого (реакции на фактор абиотического стресса) и в этих древостоях возникали очаги массового размножения непарного шелкопряда. Причем, с таким же уровнем дефолиации крон, как и в древостоях, растущих на бедных почвах.
Таким образом, согласно полученным нами результатам, механизм вспышки был обусловлен, не тонкими различиями в химическом составе листьев, не различиями в химическом составе почв, а всего лишь – в скорости реакции этих разных по почвенным условиям древостоев.
Следовательно, совершенно очевидно, что механизм возникновения вспышек массового размножения у непарного шелкопряда тесно связан с возникновением факторов абиотического стресса (периодических весенне-летних засух) и реакцией на этот фактор стресса древостоев. Древостои подвергались сильной дефолиации только после реакции их на абиотический стресс.
Таким образом, было убедительно показано, что механизм вспышек массового размножения обусловлен временным, а не постоянным ослаблением древостоев под воздействием фактора абиотического стресса. Одновременно было также очевидно, что резкий рост выживаемости популяции непарного шелкопряда происходил только после регистрации реакции на фактор абиотического стресса древостоев (Колтунов, 1998, 2006). До реакции выживаемость оставалась на низком уровне. При этом чрезвычайно важным моментом для понимания механизма возникновения вспышек было то, что рост выживаемости происходил не у всех деревьев, подвергнутых воздействию фактора абиотического стресса, а только у части деревьев. Та часть деревьев, которая осталась высоко резистентной после воздействия фактора абиотического стресса, хотя тоже подвергалась попыткам дефолиации со стороны непарного шелкопряда, тем не менее, несмотря на одинаковую начальную плотность заселения деревьев гусеницами, выживаемость их на этих деревьях была очень низкой (Колтунов, 1998, 2006). Соответственно, степень дефолиации крон также оставалась низкой.
Убедительно показана тесная взаимосвязь интенсивности засух и общей площади очагов массового размножения непарного шелкопряда и шелкопряда-монашенки (Колтунов, 1998, 2006). По нашему мнению механизм был обусловлен тем, что более интенсивные засухи вызывали и более сильную реакцию древостоев на абиотический стресс. Это, по нашему мнению, сопровождается временным резким снижением уровня энтоморезистентности древостоев. При этом, чем выше уровень реакции на фактор абиотического стресса, тем выше уровень временного снижения энтоморезистентности. Совершенно понятно, что в условиях ежегодного воздействия на древостои фактора абиотического стресса в период вспышки массового размножения (Колтунов, 2006), уровень временного ослабления древостоев, вероятно, возрастает. Аргументом в пользу этого предположения может быть то, что в условиях вспышки массового размножения непарного шелкопряда в 50 –е годы, сопровождавшейся особенно интенсивными и продолжительными засухами, как показано нами ранее (Колтунов, 1998), ослабление древостоев было настолько сильным, что уровень отпада был значительно выше, чем в последующие вспышки (Распопов, 1961, Новоженов, 1961).
Сформулирована также основная гипотеза, обусловленная механизмом воздействия фактора абиотического стресса. Она заключалась в том, что, помимо стрессового воздействия на часть древостоев, засуха вызывает, кроме временного снижения уровня энтоморезистентности еще и временные изменения физиолого-биохимических свойств листьев, которые в этот период становятся более пригодными для насекомых-филлофагов. Это сопровождается резким возрастанием выживаемости популяции непарного шелкопряда.
Было понятно, что последующие исследования необходимо проводить именно в этом направлении, так как это могло дать адекватное объяснение механизма вспышек массового размножения. И такие исследования были проведены. Результаты исследований изложены ниже.
Исходя из этого здесь для проверки этой гипотезы было бы очень важно изучить: роль количественного уровня реакции на абиотический стресс (засуху) как возможного ключевого фактора, контролирующего степень временного снижения энтоморезистентности древостоев в период засухи.
Для проверки этой гипотезы наиболее удобной моделью мы сочли изучение степени падения годичного радиального прироста в год наступления засухи как возможного ключевого критерия, показывающего уровень стрессовой реакции древостоев на засуху. Именно количественные различия в уровне реакции древостоя на стресс (засуху), в различных лесорастительных условиях и могут быть ключевым фактором, включающим каскадные изменения биохимического состава корма и его кормовую ценность. Последняя, опосредованно, через различия в выживаемости популяции и обеспечивает либо быстрый рост плотности, который и реализует вспышку, либо затух
Исследования проводили в очагах массового размножения зауральской популяции непарного шелкопряда в березовых лесах Челябинской, Свердловской, Курганской области, значительно нарушенных антропогенной дигрессией (рекреационной и пастбищной) (Колтунов, 1993, 1996, 1998, 2006).
Для этого в очагах закладывались постоянные и временные площади, размером 20х20 м, в количестве 60. Изучались лесотаксационные параметры почв, влажность почв, содержание в них N,P, Ca, рН почв и другие. Определяли также степень антропогенной дигрессии по стадиям (Колтунов, 1993 ).
В очагах с разной степенью дефолиации крон отбирались керны буравом Престлера (10-30%; 50-60%; 80-100%). Контролем служили древостои, недефолиированные непарным шелкопрядом.
Керны сушили. Затем с помощью микроскопа и микрометрической линейки проводили измерение годичного радиального прироста у березы.
Количественный уровень стрессовой реакции древостоя определяли по величине падения годичного радиального прироста в год наступления засухи (Хn+1/Xn). Для этого показатели годичного радиального прироста в течение трех-четырех лет, предшествующие засухе - (среднее значение) сравнивали с показателями радиального прироста в год наступления засухи (в мм).
В сходных лесорастительных условиях формировали выборку из пробных площадей со сходным уровнем дефолиации крон. Общее количество кернов в одной выборке составляло 40 шт. В составе выборки были деревья диаметром 20-25 см, одинакового возраста.
Полученные данные обрабатывали статистически с помощью F критерия Фишера (Рокицкий, 1973).
Таблица 4.1 Влияние уровня реакции березы на фактор абиотического стресса (весенне-
летние засухи) на уровень энтоморезистености
|
Наименование
вариантов
|
Изменение годичного радиального прироста березы в год засухи
(xn+1 / xn ) (x ± v)
|
F критерий
Фишера
|
Кол-во
деревьев в одном варианте
|
|
|
|
|
|
|
1. Контроль. Березовые древостои не дефолиировались в период вспышки
|
1,050± 0,0300
|
1/2 - 9,74
1/3 – 576,6
2/3 – 59,75
|
40
|
|
|
|
|
|
|
2. Дефолиация 20-30% на следующий год после наступления засухи
|
1,449± 0,0937
|
--
|
40
|
|
|
|
|
|
|
3. Дефолиация 80-100% на следующий год после наступления засухи
|
2,818± 0,7210
|
--
|
40
|
На уровне локальных очагов дендрохронологический анализ березы и сосны в очагах непарного шелкопряда и шелкопряда-монашенки на Урале однозначно показал, что за последние 45-50 лет все очаги с заметной дефолиацией крон были связаны с воздействием абиотического стресса (весенне-летних засух), что сопровождалось резким падением годичного радиального прироста (Колтунов, 1996, Колтунов, Пономарев, Федоренко, 1998). Обычно воздействие стрессового фактора и сильной дефолиации крон либо было синхронным (в тот же год), но чаще либо разделено 1-2 годами.
Как показали результаты, древостои не подвергшиеся дефолиации, наиболее слабо реагируют на абиотический стресс (засуху), или совсем не реагировали на этот фактор (табл. 4.1).
Слабо дефолиируемые березняки в сходных лесорастительных условиях более значительно реагирует на стресс, в среднем на 44,9 % - падение годичного радиального прироста.
Наиболее значительная реакция на абиотический стресс наблюдалась в березовых древостоях с очень низкой энтоморезистентностью (при дефолиации 80-100 %).
Средний уровень падения прироста составил 181,8 % от прироста в предыдущие 3-4 года (табл. 4.1 ).
Таким образом, как показали результаты исследований, уровень стрессовой реакции древостоев в очагах достаточно точно отражает степень временного снижения энтоморезистентности, которая тесно коррелирует с выживаемостью гусениц и степенью дефолиации крон (табл. 4.1).
Вероятно более сильная стрессовая реакция части древостоев, потерявших энтоморезистентность, вызывает и более значительные физико-биохимические сдвиги в этих древостоях, которые сопровождаются возрастанием их кормовой ценности и - выживаемости гусениц.
В связи с тем, что ранее нами была выявлена реакция торможения отклика древостоя на один и тот же уровень абиотического стресса (засуху), в зависимости от физико-химического состава почв, мы одновременно проверили, не происходит ли в низкорезистентных древостоях только временная задержка реакции на стресс, в результате чего подсчет в год засухи мог привести к ошибке? Исследования показали, что скорость наступления стрессовой реакции не изменяется и в год наступления засухи торможение прироста было наиболее интенсивными.
Наступление засухи совпадало с продромальной фазой вспышки. На следующий год это сопровождалось в низкорезистентных древостоях сильной или полной дефолиацией крон. В год наступления засухи в обследованных очагах, преимущественно имело место слабая и средняя дефолиация. Сильная дефолиация крон наблюдалась лишь в единичных древостоях.
Следует также отметить, что в древостоях на бедных почвах с низкой рН уровень реакции на стресс был в целом несколько ниже, чем на богатых, серых лесных почвах и черноземах с более высоким рН. В сильно нарушенных антропогенной дигрессией березняках также наблюдается тенденция к снижению уровня реакции на стресс (на 4 стадии) по сравнению с мало нарушенными древостоями.
Это, по нашему мнению, отражает более высокую адаптацию к стрессовым факторам древостоев, растущих на бедных почвах за счет общего снижения интенсивности реакции на различный стресс (как засуху, так и дефолиацию), как показано нами ранее (Колтунов, 1993, 1996, 2006). По этой причине высокобонитетные древостои березы на северной границе ареала непарного шелкопряда реагировали на наступление засухи заметно выше, чем низкопродуктивные древостои в южной части ареала зауральской популяции, в степной зоне. При этом следует учесть, также, что интенсивность засухи в северной части ареала этой популяции заметно ниже, чем в центре и на юге. Показано, что более сильная реакция на стресс (как засуху, так и дефолиацию) коррелирует с особенно низкой устойчивостью этих древостоев к дефолиации и вызывает частичный отпад березы в северной части ареала (Колтунов, 1993, 1996, 2006).
Результаты показали, что уровень реакции на абиотический стресс контролируется, главным образом, влажностью почв, так как в дефолиированных древостоях влажность почв была наиболее важной; в слабо дефолиированной она была меньше, а в сильно дефолиированной - самой низкой.
Это хорошо коррелирует и с различиями в геоботаническом составе травяного яруса. В контроле процент степных видов и злаков был наименьшим, в сильно дефолиированных древостоях - максимальным.
Кроме того, присутствие осины в составе древостоя также косвенно отражает степень увлажненности этих биотопов. В контроле примесь осины была наибольшей, в слабо дефолиированной - меньше, в сильно дефолиированных древостоях осина полностью отсутствовала.
Вследствие более выраженной реакции на стресс в низкорезистентных древостоях общие потери прироста древостоя от засухи и дефолиации возрастают по сравнению с чистыми потерями в высокорезистентных древостоях только от возрастания фактора засухи.
Исследования показали, что абиотический стресс является одним из ключевых факторов вспышек массовых размножений у непарного шелкопряда в аридной зоне. Убедительно показана количественная взаимосвязь уровня реакции древостоев на фактор абиотического стресса и степень временного снижения их энтоморезистентности.
Уровень реакции древостоев, нарушенных рекреационной и пастбищной дигрессией на абиотический стресс контролируется большой группой факторов, среди которых наиболее важное значение имеет влажность почв. Но он не является единственным фактором, так как возможность реализации вспышек контролируется очень широкой группой факторов различного характера (внутрипопуляционные, абиотический стресс, фитоценотические, почвенные, климатические и другие). Он не является и общим механизмом динамики численности у всех насекомых, а свойственен группе насекомых-фитофагов с широким фазовым портретом динамики численности, которые образуют периодические крупномасштабные вспышки. Вполне вероятно, что колебания численности у других групп насекомых с низкой амплитудой могут и не быть связаны с фактором абиотического стресса. Хотя подъем численности насекомых-фитофагов - членов березовой консорции также был связан с воздействием этого стресса. Можно также предполагать, что и относительно незначительные колебания численности популяции у непарного шелкопряда на невспышечном уровне могут быть не связаны однозначно с воздействием такого стрессового фактора как засухи. На локальное изменение численности популяции оказывает воздействие очень большое количество факторов.
Таким образом, механизмы, контролирующие параметры энтоморезистентности древостоев, представляют собой иерархическую систему. По нашему мнению, фактор абиотического стресса является подсистемой высокого иерархического уровня, так как именно этот фактор обеспечивает каскадное включение целой группы других факторов низшей иерархии, таких как изменение кормовых свойств листьев и хвои и др.
Следовательно фактор абиотического стресса (засухи) является ключевым фактором реализации механизмов вспышки, так как после его воздействия (за 3 года до вспышки) он инициирует подъем численности популяции непарного шелкопряда. Его роль сводится как стрессовому воздействию на древостои, индукции временной потери энтоморезистентности и реализации возможности сильной дефолиации крон, так и к опосредованному воздействию на популяцию через резкое изменение качества кормового субстрата, которое обеспечивает активацию механизма быстрой адаптации непарного шелкопряда к изменившимся условиям среды. Активация этого механизма инициирует быстрое возрастание биотического (вспышечного) потенциала у непарного шелкопряда. Как показали результаты, приведенные в главе 5, сразу после воздействия фактора абиотического стресса экологическая плотность популяции начинает возрастать, происходит быстрый подъем численности популяции.
Как показали результаты (Колтунов, 1993,2006; Колтунов и др., 1998, 2010) воздействие фактора абиотического стресса выполняет эту же функцию и в другие фазы вспышки, поддерживая биотический (вспышечный) потенциал на высоком уровне.
Нами показано, что у некоторых вспышек функцию фактора абиотического стресса непосредственно перед началом подъема численности (начало продромальной фазы) выполняют не полные весенне-летние засухи (май-июль), а неполные локальные весенние засухи (майские), которые, как уже отмечалось выше (Колтунов, 2006), более важны для выживания популяции, чем позднелетние и осенние.
Как установлено нами (глава), а, также, показано другими авторами (Бенкевич, 1962) дополнительно эту роль выполняют и иные абиотические факторы (отклонения зимних температур и др.).
Кроме этого, фактор абиотического стресса оказывает и прямое положительное воздействие на условия роста и развития популяции за счет оптимизации температурного режима.
При отсутствии такого фактора абиотического стресса, как засухи на следующий год, на фоне нормальных гидротермических условий, развитие вспышки способно продолжаться за счет фактора инерционности (после возрастания вспышечного потенциала он некоторое время способен сохраняться и в менее благоприятных климатических условиях. Возрастание вспышечного потенциала происходит за счет быстрой адаптации эруптивных видов насекомых к стабильно благоприятным условиям среды (климатические условия и изменение качества кормового субстрата).
Возросший вспышечный потенциал позволяет преодолеть кратковременные неблагоприятные условия среды. При продолжении сохранения неблагоприятных условий среды (высокая влажность) более, чем в течение одного года, как правило, вспышка начнет затухать, так как включается механизм адаптации уже к изменившимся условиям среды обитания. Особенно, если высокий гидротермический коэффициент наблюдается в мае (Колтунов,2006; Колтунов и др., 2010).
При этом, как показано нами ранее (Колтунов, 1993, 2006;Колтунов и др., 1998, 2010), популяция непарного шелкопряда в северной части ареала зауральской популяции (Свердловская обл.) достаточно хорошо адаптировалась к реализации вспышки в менее благоприятных климатических условиях и может протекать в условиях гидротермических коэффициентов, близких к нормальным, или даже повышенным. Но, при этом, обязательным условиям для реализации вспышки, остается синхронизация начала подъема численности популяции с весенними засухами и зимними низкими температурами, которые являются ключевыми абиотическими факторами, инициирующими включение биотического механизма быстрой адаптации к изменению условий обитания и роста вспышечного потенциала.
Для более подробного ознакомления с этой темой рекомендуем следующие монографии:
Колтунов Е.В. Насекомые-фитофаги лесных биогеоценозов в условиях антропогенного воздействия.
Екатеринбург, Изд-во «Наука», 1993, 137 с
Колтунов Е.В., Пономарев В.И., Федоренко С.И. Экология непарного шелкопряда в условиях
антропогенного воздействия. Екатеринбург: Изд-во УрО РАН, 1998. 214 с.
Колтунов Е.В. Экология непарного шелкопряда в лесах Евразии. Екатеринбург. Изд-во УрО РАН. 2006.
260 с. (Скачать монографию (PDF)).
Колтунов Е.В., Бахвалов С.А., Мартемьянов В.В. Факторы и экологические механизмы популяционной
динамики лесных насекомых-филлофагов. Новосибирск: Изд-во СО РАН, 2010 г., 300 c.
ISBN 978-57692-1149-2 (Скачать монографию (PDF)).