Сибирский шелкопряд (Dendrolimus superans sibiricus Tschetv.)
Сибирский шелкопряд (Dendrolimus superans sibiricus Tschetv.)
Сибирский шелкопряд ( Dendrolimus superans sibiricus Tscetv.) в азиатской части России является одним из наиболее опасных насекомых-вредителей хвойных лесов, особенно в Сибири и на Дальнем Востоке. Периодические крупномасштабные вспышки массового размножения этого фитофага приводят к значительным изменениям структуры таежных лесов, разрушению древостоев и смене лесных формаций.
Очаги массового размножения отмечаются ежегодно на площади от 4,2 тысяч до 6,9 миллиона га (в среднем 0,8 миллиона га) и наносят существенный урон лесному хозяйству. Поэтому спутниковый мониторинг как часть энтомологического мониторинга лесов является важным элементом контроля состояния лесного покрова, обеспечивающий при надлежащем исполнении сохранение важнейших экологических функций лесов.
В России огромный вклад в разработку и реализацию биологических методов борьбы с очагами массового размножения сибирского шелкопряда внес д.б.н., проф. Талалаев Е.В. В середине 1990-х годов от шелкопряда пострадали обширные лесные насаждения в Западной и Восточной Сибири, а также на Дальнем Востоке. Только в Красноярском крае в течение четырех лет вспышка охватила территории 15 лесхозов, площадь поврежденных участков тайги составила более 600 тысяч гектаров. Уничтожено большое количество ценных кедровых насаждений. За последние 100 лет на территории Красноярского края зарегистрировано 9 вспышек вредителя. В результате были повреждены леса на площади более 10 миллионов гектаров. Применение современных инсектицидных пиретроидных и бактериальных препаратов позволило частично локализовать очаги вредителя и остановить его дальнейшее распространение.
Вместе с тем сохраняется опасность нового массового размножения сибирского шелкопряда.
Вместе с тем сохраняется опасность нового массового размножения сибирского шелкопряда.
В период между вспышками шелкопряд живет в резервациях - участках с наиболее благоприятными условиями развития. В зоне темнохвойной тайги резервации располагаются в спелых, достаточно продуктивных (II-III класс бонитета) насаждениях разнотравно-зеленомошных типов леса с участием пихты до б единиц и более, с полнотой 0.3-0.6.

Имаго сибирского шелкопряда. Фото: Natalia Kirichenko, Bugwood.org

Имаго сибирского шелкопряда.Фото: (http://www.forest.protect.ru)
Сибирский шелкопряд – крупная бабочка с размахом крыльев у самки 60–80 мм, у самца 40–60 мм. Окраска варьирует от светло-желтовато-коричневого или светло-серого до почти черного цвета. Передние крылья пересекаются тремя более темными полосами. В середине каждого крыла находится большое белое пятно, задние крылья одноцветные.
Самки откладывают яйца на хвою, преимущественно в нижней части кроны, а в периоды очень большой численности – на сухие ветки, лишайники, травяной покров, лесную подстилку. В одной кладке обычно бывает несколько десятков яиц (до 200 шт.), а всего самка может отложить до 800 яиц, однако чаще всего плодовитость не превышает 200–300 яиц.
Яйца почти шаровидной формы, в диаметре до 2 мм, сначала голубовато-зеленого цвета с темно-коричневой точкой на одном конце, затем сероватые. Развитие яиц длится 13–15 суток, иногда 20–22 суток.

Различные вариации окраски гусениц сибирского шелкопряда (фото: http://protect.forest.ru)
Гусеницы сибирского шелкопряда имеют различную окраску. Она варьирует от серо-бурой до темно-коричневой. Длина тела гусеницы 55–70 мм, на 2-м и 3-м сегментах тела имеют черные с синеватым отливом поперечные полосы, а на 4–120-м сегментах – черные подковообразные пятна (рис. ).
Первая линька происходит через 9–12 суток, вторая через 3–4. В первом возрасте гусеницы объедают только края хвоинок, во втором возрасте они объедают хвою целиком. В конце сентября гусеницы зарываются в подстилку, где зимуют под моховым покровом.
В конце апреля гусеницы поднимаются в кроны деревьев и начинают питаться, поедая хвоинки целиком, а при недостатке пищи – кору тонких побегов и молодые шишки. Примерно через месяц гусеницы линяют в третий раз, а во второй половине июля – еще раз. Осенью они уходят на вторую зимовку. В мае-июне следующего года взрослые гусеницы интенсивно питаются, причиняя наибольший вред. В этот период они съедают 95% пищи, необходимой для полного развития. Они линяют 5–7 раз и соответственно проходят 6–8 возрастов.
Гусеницы питаются хвоей почти всех хвойных пород. Но предпочитают пихту, ель, лиственницу. В меньшей степени повреждаются кедр, еще меньше - сосна. В июне гусеницы окукливаются, перед окукливанием гусеница плетет буро-серый продолговатый кокон. Куколка, длиной 25–45 мм, коричневато-красная, затем темно-коричневая, почти черная. Развитие куколки зависит от температуры и длится около месяца. Массовый лет бабочек происходит во второй декаде июля. На южных склонах гор он проходит раньше, на северных – позднее.
Цикл развития сибирского шелкопряда обычно длится 2 года. Но на юге ареала развитие заканчивается почти всегда за один год, а на севере и в высокогорных лесах иногда бывает и трехлетняя генерация. Лёт бабочек начинается во второй половине июля и продолжается около месяца. Бабочки не питаются. Размах крыльев самок составляет от б до 10 см самцов - 4-5 см. В отличие от самок, самцы имеют перистые усики. Самка откладывает в среднем около 300 яиц, размещая их по одному или группами на хвоинках в верхней части кроны. Во второй половине августа из яиц выходят гусеницы первого возраста, питаются зеленой хвоей и во втором или третьем .возрасте в конце сентября уходят на зимовку. Зимуют гусеницы в подстилке под покровом мха и слоем опавшей хвои. Подъем в крону отмечается в мае после схода снега. Гусеницы питаются до следующей осени и уходят на вторую зимовку в пятом - шестом возрасте. Весной они вновь поднимаются в кроны и после активного питания в июне плетут плотный серый кокон, внутри которого затем окукливаются. Развитие шелкопряда в куколке длится 3-4 недели.
В период между вспышками шелкопряд не наносит серьезных повреждений: его количество составляет 1-2 гусеницы на дерево, причем гусениц можно обнаружить не на каждом дереве. Численность шелкопряда регулируется различными видами паразитов: наездниками рогасом и апантелесом, мухой тахиной. Однако основными природными регуляторами численности шелкопряда в Восточной Сибири являются яйцееды теленомусы. В лиственничных лесах на количество теленомуса в большой степени влияют низовые пожары, уничтожающие яйцеедов. По этой причине очаги массового размножения шелкопряда в лиственничниках часто возникают после пожаров.
В темнохвойной тайге очаги шелкопряда образуются после нескольких лет жаркой сухой погоды летом. В этом случае гусеницы уходят на зимовку позже, в третьем либо четвертом возрасте, и превращаются в бабочек следующим летом, переходя на однолетний цикл развития. Ускорение развития гусениц является условием формирования очагов сибирского шелкопряда.

Участок хвойного леса после дефолиации сибирским шелкопрядом. (Фото Д.Л.Гродницкого).

Участок леса, дефолиированный сибирским шелкопрядом (фото: http://molbiol.ru)
Учет зимующих гусениц в подстилке проводят в октябре или начале мая Численность гусениц в кроне определяют методом околота на матерчатые полога в начале июня и в конце августа.
Возраст гусениц устанавливают по таблице, производя замер ширины головы.
Возраст гусениц устанавливают по таблице, производя замер ширины головы.
Следует иметь ввиду, что в условиях Северной Евразии леса, погибшие от шелкопряда, плохо восстанавливаются. Гусеницы уничтожают подрост вместе с древостоем, и только спустя десятилетие возможно появление малочисленного подроста лиственных пород. В старых очагах хвойные появляются лишь через 30-40 лет после усыхания древостоев, причем не везде и не всегда.
Основной причиной отсутствия естественного возобновления в шелкопрядниках является резкая экологическая трансформация растительных сообществ. Во время массового размножения шелкопряда за 3-4 недели в подстилку и почву поступает до 30 т/га объеденных фрагментов хвои, экскрементов и трупов гусениц. Буквально в течение одного сезона вся хвоя в насаждении перерабатывается гусеницами и поступает в почву. Этот опад содержит значительное количество органических веществ - благоприятного корма для почвенных бактерий и грибов, деятельность которых значительно активизируется после массового размножения шелкопряда.
Этому также способствует увеличение температуры и влажности почвы, поскольку ни солнечный свет, ни осадки более не задерживаются кронами деревьев. Фактически, массовое размножение шелкопряда способствует более интенсивному протеканию биологического круговорота в результате быстрого освобождения значительных количеств вещества и энергии, заключенных в лесной подстилке.
Почва в шелкопрядниках становится более плодородной. На ней бурно развивается светолюбивый травяной покров и подлесок, происходит интенсивное задернение и часто - заболачивание. Вследствие этого сильно нарушенные насаждения сменяются нелесными экосистемами. Поэтому восстановление насаждений, близких к исходным, затягивается на неопределенный срок, но не менее чем на 200 лет (Солдатов и др., 2000).
Вспышки массового размножения сибирского шелкопряда в лесах УрФО
В целом, несмотря на большое количество работ по экологии сибирского шелкопряда в 50-60 годах, многие особенности экологии зауральской популяции в условиях глобального антропогенного воздействия остаются неизученными.
Вспышки массового размножения сибирского шелкопряда в лиственничных лесах Предуралья отмечаются с 1900 г. [Ханисламов, Яфаева, 1962], В темнохвойных равнинных лесах Зауралья в Свердловской и Тюменской областях предыдущая вспышка наблюдалась в 1955-1957 г.г, а следующая - в 1988-1992 г.г. Первая вспышка в лесах Свердловской области обнаружена в 1955 г. на территории Тавдинского и Туринского лесхозов. Общая площадь очагов составила, соответственно, 21.000 га и 1600 га. На территории Тавдинского лесхоза крупные очаги образовались и раньше. Обращает внимание, что указанные лесхозы в течение многих десятилетий были местом интенсивных заготовок древесины. Поэтому хвойные леса подверглись антропогенной трансформации и в настоящее время имеют примесь вторичного березняка с сосной, елью и пихтой в подросте. Следует отметить, что новая вспышка (1988-1992 г.г.) в Свердловской области зарегистрирована в других лесхозах. В наибольшей степени она сформировалась в лесах Таборинского района. Общая площадь очагов составила 862 га, отдельные очаги наблюдались и при авианаблюдении в Гаринском районе.
Исследования показали, что на 50% площадей, затронутых очагами в 1988-1992 г.г., основной лесообразующей породой является береза с пихтой и елью в составе подроста (Колтунов, 1996, Колтунов и др., 1997). Пихтовый подрост сильно дефолиировался сибирским шелкопрядом и, в основном, усох. В результате нанесен существенный ущерб развитию хвойного хозяйства в этих лесхозах. Первичные очаги массового размножения сибирского шелкопряда возникли в 1988 г. в насаждениях с пихтовым подростом. В 1993 г. вспышка полностью затухла. На территории ХМАО-ЮГРЫ вспышка массового размножения затухла в 1992 г. В отдельных кварталах наблюдалась дефолиация сибирским шелкопрядом ели, в результате чего она также быстро усыхала. Как показали обследования в очагах этого фитофага в период вспышки, развитие зауральской популяции происходит, преимущественно, по двухгодичному циклу. В целом исследования показали, что топография очагов широкого шелкопряда в хвойных лесах Свердловской области совпадает с участками леса, нарушенными антропогенным воздействием.
На территории Ханты-Мансийского Автономного округа вспышка массового размножения сибирского шелкопряда обнаружена на территориях Междуреченского, Урайского, Тобольского, Вагайского и Дубровинского лесхозов. Общая площадь очагов составила 53000 га. Мы проводили наиболее детальные исследования в очагах массового размножения сибирского шелкопряда в Междуреченском лесхозе.
На территории Южно-Кондинского ЛПХ в течение последних 20 лет происходила наиболее интенсивная промышленная лесозаготовка. Как показали результаты пространственная структура очагов массового размножения сибирского шелкопряда в этом лесхозе однозначно не совпадает с лесами, подвергнутыми наиболее интенсивному антропогенному воздействию (в первую очередь - вырубке). Наиболее крупные очаги (в западной части лесхоза) совершенно не затронуты антропогенным воздействием. Рубки в лесах до вспышки не проводились. Никаких других типов антропогенного воздействия мы также не обнаружили. Анализ лесотаксационных параметров древостоев в этой группе очагов показал, что эти леса имеют обычную для данного типа лесорастительных условий продуктивность и не являются ослабленными. В то же время рядом с другими, более мелкими очагами наблюдаются вырубки, в отдельных случаях - пожары. Некоторые из очагов с сильной дефолиацией крон древостоев ранее пройдены рубками.
Как показали результаты антропогенное воздействие в темнохвойных равнинных лесах Зауралья не является ключевым фактором формированияочагов массового размножения сибирского шелкопряда, хотя вклад его несомненен. В условиях умеренного антропогенного воздействия основным фактором организации пространственной структуры очагов являются лесорастительные условия в экотопах и особенности микрорельефа. Так наиболее крупные очаги примыкают к руслам рек и местам с микроповышениями, что известно ранее [Коломиец, 1960,1962; Ивлиев, 1960]. Особенно важным фактом является то, что леса в районах очагов не были заметно ослаблены под влиянием антропогенных факторов. Уровень антропогенной трансформации этих лесов был крайне незначителен, не выше 1 стадии в отдельных экотопах (5-10% лесов). Как показал геоботанический анализ травяного яруса, травяной покров в этих лесах не изменен.
Таким образом, в наибольшей степени на этих лесах сказывается лишь соседство с вырубками (изменение светового и ветрового режима) и в меньшей степени – рубки, проведенные несколько десятилетий назад в отдельных из них.
Анализ радиального прироста деревьев в очагах и за их границами подтверждает наш вывод о сохранении устойчивости лесов в целом, которые подверглись дефолиации. Пониженный радиальный прирост деревьев в очагах мы связываем с адаптивной реакцией древостоев на лесорастительные| условия, но не с их ослабленностью, так как эти различия мы обнаруживали не в последние годы, а на протяжении 50 и более лет.
Характерной чертой особенностей динамики дефолиации древостоев в период вспышки в равнинных лесах Зауралья было явное предпочтение дефолиации пихты в подросте в начале вспышки, затем - пихты в основном ярусе, позже - ели и кедра. Сосна дефолиировалась очень слабо. Поэтому в чистых сосняках очаги не формировались. Исследование зауральской популяции сибирского шелкопряда в очагах показало, что в эруптивной фазе и перед затуханием вспышки отрождаемость имаго была очень низкой и колебалась от 2 до 30%, составляя в среднем 9,16%.
Большая часть популяции куколок погибает. Наиболее значительный процент популяции гибнет от инфекционных болезней (бактериозы и вирус гранулеза). Гибель от этих причин составляет от 29,0 до 64,0%, в среднем, 47,7%. Бактериальные инфекции составляли основной процент из причин гибели от этой группы болезней. Вирусные инфекции встречались значительно меньше. Следует также отметить, что микроскопический анализ погибших гусениц в очагах как в Свердловской, так и в ХМАО убедительно показал, что затухание вспышек не сопровождалось вирусной эпизоотией (вируса гранулеза).
Второй по важности причиной гибели популяции куколок в этот период вспышки была гибель от неидентифицированных причин, не связанных с инфекционными болезнями (18-29%, в среднем: 25,0%). Смертность популяции от паразитов занимает самое низкое место в ранговой структуре основных факторов смертности. Она в этот период колеблется от 5 до 22,0%, в среднем: 12,67%.
Полученные нами результаты хорошо согласуются с данными других исследователей по другим популяциям сибирского шелкопряда [Ханисламов, Яфаева, 1958; Болдаруев, 1960,1968; Ивлиев, 1960; Рожков, 1965].
В период затухания вспышки массового размножения сибирского шелкопряда в лесах Ханты-Мансийского АО в подстилке обнаруживались до 30 гусениц на 1м2 , погибших от инфекционных болезней.
Как показал корреляционный анализ основных популяционных параметров, выживаемость гусениц в кроне обусловлена, преимущественно, уровнем энтоморезистентности древостоев и возрастание энтоморезистентности тесно коррелирует со снижением выживаемости гусениц и плотностью их популяции в кроне (г=0,732). При возрастании выживаемости гусениц в кроне и увеличении степени дефолиации одновременно регистрируется возрастание гибели куколок от инфекционных болезней (r=0,541), но при этом одновременно резко снижается процент паразитированных куколок (г=-0,973). Последнее обстоятельство вполне объяснимо с точки зрения запаздывания динамики паразита от хозяина при высокой плотности последнего [Исаев и др., 1984].
Как показали результаты интересной особенностью древостоев, усохших после дефолиации сибирским шелкопрядом в равнинных темнохвойных лесах Ханты-Мансийского АО, было, практически полное, отсутствие заселения их насекомыми-ксилофагами в течение 1-2 лет после усыхания, хотя в неповрежденных сибирским шелкопрядом лесах наблюдается заселение ксилофагами усыхающих древостоев и отдельных деревьев.
При этом следует отметить, что запас ксилофагов в районах очагов достаточный. Кроме того, на вахтовых участках и на складах запаса в Южно-Кондинском ЛПХ оставленные без обработки хлысты быстро заселяются насекомыми-ксилофагами. Мы связываем замедление заселения ксилофагами усохших древостоев после дефолиации их сибирским шелкопрядом в большей степени с повышенной влажностью древесины. Это, по нашему мнению, было обусловлено активным транспортом воды корневой системой деревьев после дефолиации крон на фоне прекращения транспирации вследствие отсутствия хвои.
Исследования в очагах массового размножения сибирского шелкопряда в Зауралье показали: последняя вспышка этого фитофага в темнохвойных лесах равнинного Зауралья наблюдалась 33 года назад. Можно предполагать, что цикличность вспышек этого фитофага на западной границе ареала тесно связана с периодичностью наиболее сильных засух 1955 и 1986 г.г.. Наиболее сильная засуха (в 1955 г.) сопровождалась и большей площадью очагов этого фитофага в Зауралье.
Ранее в Кондинском лесхозе вспышек сибирского шелкопряда не было. Дендрохронологический анализ кернов пихты и ели (за последние 100-120 лет), проведенный нами, показал, что древостои как в очаге, так и за его границами, ранее не подвергались заметной дефолиации. На основании полученных нами результатов можно предполагать, что происходит постепенное проникновение сибирского шелкопряда на север и возникновение в этих местообитаниях вспышек массового размножения, которые там ранее не наблюдались. Вероятно, это обусловлено постепенным потеплением климата.
Взаимосвязь пространственной структуры очагов и антропогенного воздействия на лесные биогеоценозы убедительно не прослеживается. Очаги выявлены как в участках леса, где велись активные рубки, так и в совершенно незатронутых рубками лесах, которые значительно удалены от дорог, зимников и поселков.
На основании полученных результатов установлено, что в условиях антропогенной трансформации темнохвойных лесов Зауралья наиболее крупные очаги сибирского шелкопряда способны возникать как в совершенно ненарушенных лесах, так и в лесах, подверженных воздействию антропогенных факторов.
Сравнительный анализ пространственно-временной структуры очагов в течение двух последних вспышек показывает, что очаги массового размножения каждый раз образуются в разных экотопах и пространственно совершенно не совпадают. Как показали результаты исследований, первые очаги в каждом из обследуемых лесхозов возникли в 1988 г. одновременно с другими очагами в более южных районах Тюменской области. Это исключает возможность возникновения их миграционным путем из южной части ареала. Вероятно, популяция в фазе депрессии была и в северной части ареала этой популяции.
На западной границе ареала этого фитофага вспышки имеют быстротекущий характер. Это хорошо объяснимо узостью временного интервала климатического оптимума в период засухи. Учитывая это, а также наличие двухгодичного цикла у гусениц сибирского шелкопряда это дает хорошие перспективы снижения экономического ущерба от вспышек за счет применения активных мероприятий в период, непосредственно перед эруптивной фазой вспышки. Сохранение высокого потенциала вспышки возможно лишь в этот узкий период засухи. Поэтому обработка очагов в этот период исключит вероятность образования крупных повторных шагов.
Как показали результаты сравнительного анализа лесотаксационных параметров 50 пробных площадей, заложенных в очагах массового размножения зауральской популяции сибирского шелкопряда в Таборинск|ом лесхозе Свердловской области, очаги формировались в древостоях с различной полнотой: от 0,5 до 1,0, в среднем - 0,8 (табл. 3.1,3.2). Корреляционный анализ показал, что площади очагов положительно коррелировали с классом бонитета (R=0,541) (с худшими условиями роста), средней высотой (R=0,54) и отрицательно коррелировали с полнотой (R=-0,54).
Тем не менее, обращает внимание, что из 50 пробных площадей только у 36 % площадей с полнотой, ниже, чем 0,8 формировались очаги массового размножения зауральской популяции сибирского шелкопряда, тогда как в подавляющем большинстве пробных площадей была полнота 0,8 и выше. Средний уровень дефолиации более низкополнотных древостоев составляет, в среднем, 54,5 %, тогда как высокополнотных (с полнотой, равной 0,8 и более) -70,1 %, но различия были статистически недостоверны. Вероятно, это свидетельствует о том, что на уровень дефолиации оказывает влияние комплекс других факторов, который является общим для группы древостоев. Вклад этой группы факторов в уровень энтоморезистентности древостоев был значительно выше, чем влияние полноты древостоев.
Исследования показали, что этим фактором являются почвенно-эдафические условия в экотопах. Так все древостои на пробных площадях, которые находились на гривах, в более сухих местообитаниях, были дефолиированы наиболее сильно, по сравнению с древостоями на равнинных частях рельефа, или микропонижениях. Корреляционный анализ степени дефолиации с другими лесотаксационными параметрами также не выявил статистически достойной ее взаимосвязи с классом бонитета (г=0,285). Тем не менее, средний уровень дефолиации самых низкобонитетных древостоев (с классом бонитета: 4-5 А) был 45,55%, тогда как в наиболее высокобонитетных – 68,33%. Различия статистически достоверны (при Р=0,01). Отсутствие достоверной линейной корреляционной зависимости также было обусловлено, вероятно, сильным доминированием фактора почвенно-эдафических условий. Это сопровождается сильной дефолиацией древостоев, значительно различающихся по классу бонитета. Нельзя исключить также возможного влияния фактора локальной миграции гусениц из полностью дефолиированных высокобонитетных древостоев в расположенные рядом низкобонитетные. Хотя следует отметить, что гусеницы в кроне регистрировались нами в обоих группах древостоев. Следовательно локальная миграция в любом случае не была основной причиной сильной дефолиации низкобонитетных древостоев.
Анализ результатов показывает, что в условиях равнинных темнохвойных лесов Свердловской обл. наблюдается определенная тенденция к преимущественному формированию очагов с наиболее сильной дефолиацией крон в древостоях с более высоким классом бонитета. Но заметного избегания низкобонитетных древостоев также не происходит. Очаги с разной степенью дефолиации крон возникают в древостоях с разным классом бонитета. Но наиболее низкая энтоморезистентность и сильная дефолиация характерна для насаждений с наиболее высоким классом бонитета. Учитывая тесную взаимосвязь степени дефолиации с уровнем энтоморезистентности древостоев при одинаковой исходной плотности популяции можно предполагать, в данных лесорастительных условиях в результате воздействия фактора абиотического стресса (засухи) энтоморезистентность древостоев с более высоким классом бонитета снижается больше, чем низкобонитетных древостоев, что сопровождается более высокой дефолиацией крон высокобонитетных древостоев.
Анализ особенностей состава древостоев в очагах массового размножения сибирского шелкопряда в Свердловской области позволил идентифицировать два основных типа стратегии формирования очагов по отношению к составу древостоев.
1 тип стратегии. Очаги возникают в основном ярусе леса. Эти древостои наиболее часто расположены на гривахи повышенных частях рельефа в более сухих типах леса. Очаги с наиболее значительной дефолиацией древостоев формируются в елово-пихтовых и пихтово-еловых древостоях с примесью березы (6П2Е2Б, 5Е2П2Б). В составе подроста имеется пихта, которая и подвергается сильной дефолиации в первую очередь. В очагах данного типа всегда наблюдается сильная дефолиация. Очаги, как правило, концентрированного типа с хорошо выраженной границей. Обследования в очагах показали, что в этих условиях, оптимальных для реализации вспышки, преобладающий состав пород не имеет критического значения и может варьировать в достаточно широких пределах. Тем не менее, в лесах с преобладанием пихты в основном ярусе и подросте формирование очагов с сильной дефолиацией наиболее вероятно. Можно предполагать, что в оптимальных почвенно-эдафических условиях общий уровень падения энтоморезистентности и пихты и ели выше, чем уровень различий энтоморезистентности между этими породами в менее оптимальных местообитаниях. По составу древостоя в этих очагах вообще не было насаждений с преобладанием пихты, а имеется ельник с пихтой и березняк с пихтовым подростом.
Следует отметить, что в очагах этого типа в Свердловской области наблюдается обычно быстрое заселение усохших древостоев насекомыми-ксилофагами, тогда как в очагах сибирского шелкопряда в лесах Ханты-Мансийского автономного округа, как указывалось выше, заселение погибших древостоев ксилофагами почти не происходили.
2 тип стратегии. Очаги возникают не в основном типе леса, а в подросте. Это характерно для участков леса, подвергшихся вырубкам. В таком типе леса возникновение очагов происходит вне зависимости от породного состава основного яруса. Это обусловлено тем, что во многих типах леса, подвергшихся сильной вырубке, имеется обильный подрост пихты, которая полностью дефолиируется и усыхает. Часто основным ярусом в этих типах древостоев служит береза, реже сосна и другие породы. Следовательно, данные типы леса являются промежуточными в динамике сукцессии, когда смена пород происходит чаще всего через березу [Колесников, 1961, 1973].
Как показали исследования в этих типах леса очаги формируются при более широкой амплитуде лесорастительных и почвенно-эдафических условий. Очаги этого типа часто встречаются не на повышенных, а на равнинных элементах рельефа, но не избыточно увлажненных.
В очагах с сильной дефолиацией в лесах Свердловской обл. очень редко в составе основного яруса встречается осина, так как она является индикатором увлажненных местообитаний. Тем не менее, в отдельных очагах с сильной дефолиацией она все-таки обнаруживается в небольшом количестве. Обычно это очаги, сформировавшиеся в равниной части рельефа, с отдельными понижениями. Как известно, такие древостои начинают повреждаться сибирским шелкопрядом после длительной засухи, снижающей влажность почвы (Коломиец, 1958,1962).
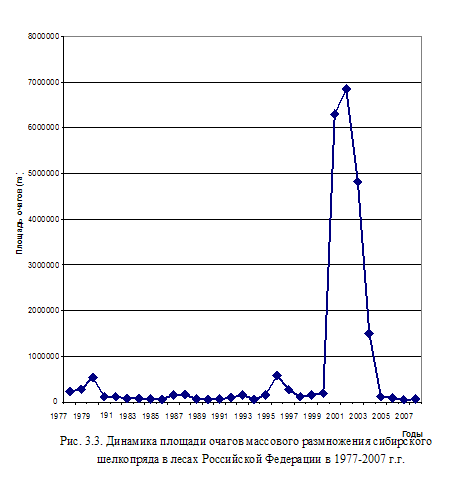
Последняя вспышка массового размножения сибирского шелкопряда возникла в 1999 г. и продолжилась до 2007 г. (Рис. 3.3). Это была самая крупная вспышка на территории России за последние 30 лет.
Основную площадь составили очаги массового размножения в Сибири и на Дальнем Востоке. В Зауралье она была, наоборот, очень слабой. В лесах Челябинской обл. площади очагов в 2006 и 2007 г.г. составили, соответственно, 116 и 115 га, в лесах Тюменской обл. на 2005 г. их общая площадь составила 200 га, в последующие 2 годы они не отмечались. В лесах Свердловской обл. она отсутствовала.
Впервые нами проведены исследования особенностей развития вспышек массового размножения в лесах Свердловской обл. и Ханты-Мансийского Автономного округа (ХМАО-ЮГРА).
В целом результаты показали очень близкое сходство лесорастительных условий предпочитаемых экотопов у зауральской и западно-сибирской популяций сибирского шелкопряда. Это обусловлено близким сходством условий местообитания этих популяций в заболоченных равнинных темнохвойных лесах.
Установлено, что в условиях антропогенной трансформации темно-хвойных лесов Зауралья сибирский шелкопряд может формировать крупные очаги как в лесах, нарушенных антропогенными факторами, так и в совершенно ненарушенных лесах. Исследования показали, что умеренный уровень антропогенной трансформации равнинных темнохвойных лесов Зауралья не является доминирующим факторов возникновения очагов. Ранг этого фактора приблизительно сходен с другими факторами предпочтения природного характера, главным из которых является микрорельеф и относительно сухие местообитания.
В западной части ареала сибирского шелкопряда вспышки имеют быстротекущий характер. Возникают, преимущественно, очаги концентрированного типа. Характер пространственной структуры первичных очагов позволяет предполагать, что они возникли немиграционным путем и сибирский шелкопряд имеется в районе вспышек и в период депрессии. Формирование очагов с сильной дефолиацией наблюдается в лесах с широким диапазоном полноты и классов бонитета в ХМАО-Югре - в пихтово-еловых лесах, в Свердловской области - в производных березняках с подростом из пихты и елово-пихтовых лесах.
Дендрохронологический анализ кернов пихты и ели (за последние 100-120 лет), проведенный нами, показал, что древостои как в очаге, так и за его границами, ранее не подвергались заметной дефолиации. Следовательно, ранее в Кондинском лесхозе ХМАО вспышек массового размножения сибирского шелкопряда не было. На основании полученных нами результатов можно предполагать, что происходит постепенное проникновение сибирского шелкопряда на север миграционным путем и возникновение в этих местообитаниях вспышек массового размножения, которые там ранее не наблюдались. Вероятно, это обусловлено постепенным потеплением климата.
Установлено, что сниженный среднегодовой радиальный прирост ели и пихты в очагах массового размножения сибирского шелкопряда не является следствием ослабления лесов в последние годы, а представляет собой норму реакции на относительно сухие условия роста на гривах и микроповышениях рельефа и различия в радиальном приросте сохраняется в течение многих десятилетий.
Несмотря на явное возрастание масштабов и уровня антропогенного воздействия на равнинные темнохвойные леса Зауралья и ХМАО-Югры частота вспышек массового размножения сибирского шелкопряда не изменилась.
Сибирский шелкопряд в Зауралье и Западной части Западной Сибири по-прежнему является очень опасным вредителем, наносящим значительный эколого-экономический ущерб лесному хозяйству региона. Поэтому считаем необходимым усилить мониторинг зауральской популяции сибирского шелкопряда.
Рекомендации по борьбе с сибирским шелкопрядом
Совершенно очевидно, что основой успешной борьбы с сибирским шелкопрядом является периодический мониторинг численности этого фитофага в резервациях. В связи с тем, что возникновение вспышек массового размножения сибирского шелкопряда тесно синхронизировано с весенне-летними засухами, надзор в этот период необходимо существенно усилить.
Рекомендуемый мониторинг только на постоянных пробных площадях и в резервациях сибирского шелкопряда для эффективного управления популяцией совершенно недостаточен.
Необходимо анализировать состояние и численность популяции и в других участках леса.
Относительно энтомофагов, необходимо иметь ввиду, что, при благоприятных климатических условиях происходит настолько интенсивный рост вспышечного (биотического) потенциала популяции, что даже очень сильное поражение яиц сибирского шелкопряда теленомусом не предотвращает возникновения вспышки массового размножения. Вместе с тем, сильная пораженность зимующих гусениц паразитами и болезнями обязательно должна учитываться при прогнозировании ожидаемой дефолиации, так как она реально значительно снижает ожидаемый уровень дефолиации крон древостоев в очаге.
Мероприятия по борьбе должны быть запланированы на период начала вспышки массового размножения, при прогнозировании более, чем 30%-ной дефолиации пихты и ели, сосны кедровой, или сильной (70%-ной) дефолиации лиственницы.
Как правило, проводится авиаобработка лесов инсектицидами. Наиболее перспективным биологическим препаратом к настоящему времени является лепидоцид.