Непарный шелкопряд (Lymantria dispar L.)
Для бабочки непарного шелкопряда характерен резкий половой диморфизм. Самец в размахе крыльев 35-40 мм, окрашен темнее, чем самка, передние крылья буровато-серые с поперечными темными волнистыми полосами. Задние крылья одноцветные бурые. Брюшко узкое, усики широкогребенчатые. Самка крупнее, в размахе крыльев 55-70 мм. Обе пары крыльев грязно-белого цвета, передние крылья с четкими темно-коричневыми зигзагообразными полосами (рис. 1). Тело самки крупное, брюшко заметно утолщенное и густо покрыто коричнево-желтыми волосками.

Рис. 1. Имаго непарного шелкопряда (фото: JohnH. Ghent, USDA Forest Service, Bugwood. org)
Усики щетинковидные, односторонне гребенчатые. Яйца вначале розоватые, позже — темно-серые, уложены в округлые кучки, покрытые светлыми желто-коричневыми волосками с тела самок (рис. 3). Эти образования, содержащие 500-1000 яиц, называются “губки”. Волоски охраняют яйца от мороза (яйца зимуют), а также делают их незаметными в окружающей среде (мимикрия).

Рис. 2. Гусеница непарного шелкопряда (фото: Колтунова Е.В.)

Рис. 3. Яйцекладки непарного шелкопряда на березе в березовых
лесах Зауралья (фото: Колтунова Е.В.)
У гусениц характерная желтоватая, с темным мраморным рисунком, довольно крупная голова с двумя коричневыми полосками. Тело цилиндрической формы. Молодые, только что вышедшие гусеницы, черные, покрыты волосками. У гусениц первого возраста на втором, третьем и десятом сегментах тела желтоватые пятнышки. Взрослые гусеницы имеют длину 40-80 мм. Основная окраска от серой до желто-коричневой с тремя тонкими желтыми линиями или с одной более широкой продольной темно-коричневой линией на спине. На первых пяти сегментах по 2 сине-фиолетовые бородавки, на всех остальных — по 2 красные бородавки (рис. 2). Куколки мумиеобразные, темно-коричневые, матовые, длиной 20-30 мм, с редкими пучками волосков и с крючочками на конце тела. Уже на куколках можно установить будущий пол, кроме иного, по половым отверстиям. В отличие от шелкопряда-монашенки волоски на спинной стороне коричнево-желтые.
Непарный шелкопряд повреждает большинство лиственных пород (более 300 видов). Гусеницы нападают чаще всего на плодовые деревья, с которых легко переходят на дуб, граб, липу и многочисленные другие породы. Вспышки массового размножения этого вредителя чаще всего происходят в южных районах в дубовых насаждениях.
Бабочки появляются в природе начиная с конца августа до сентября, а в теплых районах — на месяц раньше. Первыми начинают лёт самцы, на несколько дней раньше самок. Самки тяжелые, обычно не летают и сидят на коре стволов. Самцы, особенно в вечернее время, живо летают и разыскивают самок, к которым их привлекают, и с относительно больших расстояний, сексуальные атрактанты (феромоны). Оплодотворенные самки откладывают яйца в кучки на кору нижней части лиственных деревьев, в нормальных условиях на высоту максимально 4 м. Одна самка откладывает в среднем 250-500 яиц. Бабочки живут всего около двух недель. Перезимовавшие яйца содержат маленьких гусениц, которые выходят только в апреле следующего года, когда средняя температура достигает 10 ° С. Вначале маленькие гусеницы не питаются, живут вместе и только через несколько дней расползаются по кроне. Гусеницы первого возраста выгрызают в молодых листьях небольшие отверстия, старшие гусеницы поедают листья особым, для них характерным, способом. Часто остатки листьев опадают на землю. При недостатке пищи гусеницы поедают незрелые ткани побегов, цветы, почки. Установлено, что гусеницы средних возрастов в насаждениях с подростом и влажной лесной подстилкой часто мигрируют по деревьям вниз.
Молодые гусеницы питаются в основном днем, интенсивное питание гусениц старших возрастов (от 4 и выше) происходит в основном ночью и только в случае вспышки массового размножения — и днем. Примерно через 70-80 дней, в теплых южных областях через 45-50 дней, после усиленного питания гусеницы готовы к окукливанию. Окукливание происходит в половине июля, на юге — уже в конце июня. Стадия куколки продолжается примерно 10-15 дней.
Оптимальные условия для своего развития непарный шелкопряд находит в изреженных лесах. Опасным вредителем этот вид является особенно потому, что обычно повреждает неравномерно более широкие пространства и в течение градации, как правило, через трех- четырехлетние периоды, распространяется дальше. Периодичность градаций в средней полосе 5-8 лет. Большинство деревьев после повреждений, которые наиболее интенсивны в конце июня и в июле, восстанавливаются и вновь зеленеют. Если же в критическом периоде было очень сухо, погибают отдельные дубы, а иногда и целые дубовые насаждения. При вспышках массового размножения непарный шелкопряд встречается не только в кроне деревьев, но и в жердняках и молодняках.
Непарный шелкопряд распространен в теплых областях по всей Европе (кроме северных районов), граница распространения идет через Средиземноморье до северной Африки. Известен и в северной Монголии, в Китае (до границы палеарктической области), затем в Корее и Японии. В 19 веке был завезен из Европы в Северную Америку (штат Массачусетс), где нашел пригодные условия для своего развития и до 1952 года распространился практически по всей территории США, став там опасным вредителем.
В России распространен по всей европейской части. Северная граница проходит от линии Петербург — Волгоград — Пермь. Встречается в Крыму и на Кавказе, на Урале, на Алтае, в северном Казахстане, Средней Азии, в Сибири и на Дальнем Востоке.
На начало 2007 года очаги непарного шелкопряда действовали на площади 813,2 тыс. га в 30-ти регионах России. Основные заселенные площади располагались в Дальневосточном округе (600,5 тыс. га). Там же соответственно находились наиболее крупные очаги вредителя, которые действовали в Приморском (490,0 тыс. га) и Хабаровском (110,2 тыс. га) краях В Уральском округе непарный шелкопряд встречается, в основном, в лиственных насаждениях Курганской, Тюменской и Челябинской областей.
В лесах Свердловской области фитофаг отмечается периодически, очаги действуют на площадях, не превышающих 3,5 тыс. га. В Ханты-Мансийском и Ямало-Ненецком автономных округах вспышек численности вредителя не отмечалось за весь период наблюдений. Максимальная площадь, заселённая непарным шелкопрядом, выявлена в 1996 году в насаждениях Курганской (165,0 тыс. га) и Тюменской (260,9 тыс. га) областей. Основные повреждаемые породы региона – берёза и дуб черешчатый.
В лесах Свердловской области фитофаг отмечается периодически, очаги действуют на площадях, не превышающих 3,5 тыс. га. В Ханты-Мансийском и Ямало-Ненецком автономных округах вспышек численности вредителя не отмечалось за весь период наблюдений. Максимальная площадь, заселённая непарным шелкопрядом, выявлена в 1996 году в насаждениях Курганской (165,0 тыс. га) и Тюменской (260,9 тыс. га) областей. Основные повреждаемые породы региона – берёза и дуб черешчатый.
Динамика вспышек массового размножения непарного
шелкопряда в Зауралье
Особенности популяционной динамики непарного шелкопряда в лесах Зауралья исследовались с 50 г. годов 20 века (Распопов, 1961, 1970, 1977; Распопов, Рафес, 1978; Соколов, 1988, 1990; Колтунов и др., 1992, 1998; Колтунов, 1993, 1996, 2000, 2001).).
В лесах Зауралья зарегистрировано 5 последовательных вспышек массового размножения непарного шелкопряда в березовых лесах. В челябинской области первая вспышка массового размножения возникла в 1953-1958 г.г., затем в 1963-1969 г.г., 1973-1980 г.г., 1984- 1990 г.г., в 2003 г. возникли небольшие очаги в лесах, граничащих с Башкортостаном, где развивалась интенсивная вспышка массового размножения. На остальной территории Челябинской области вспышек не было.
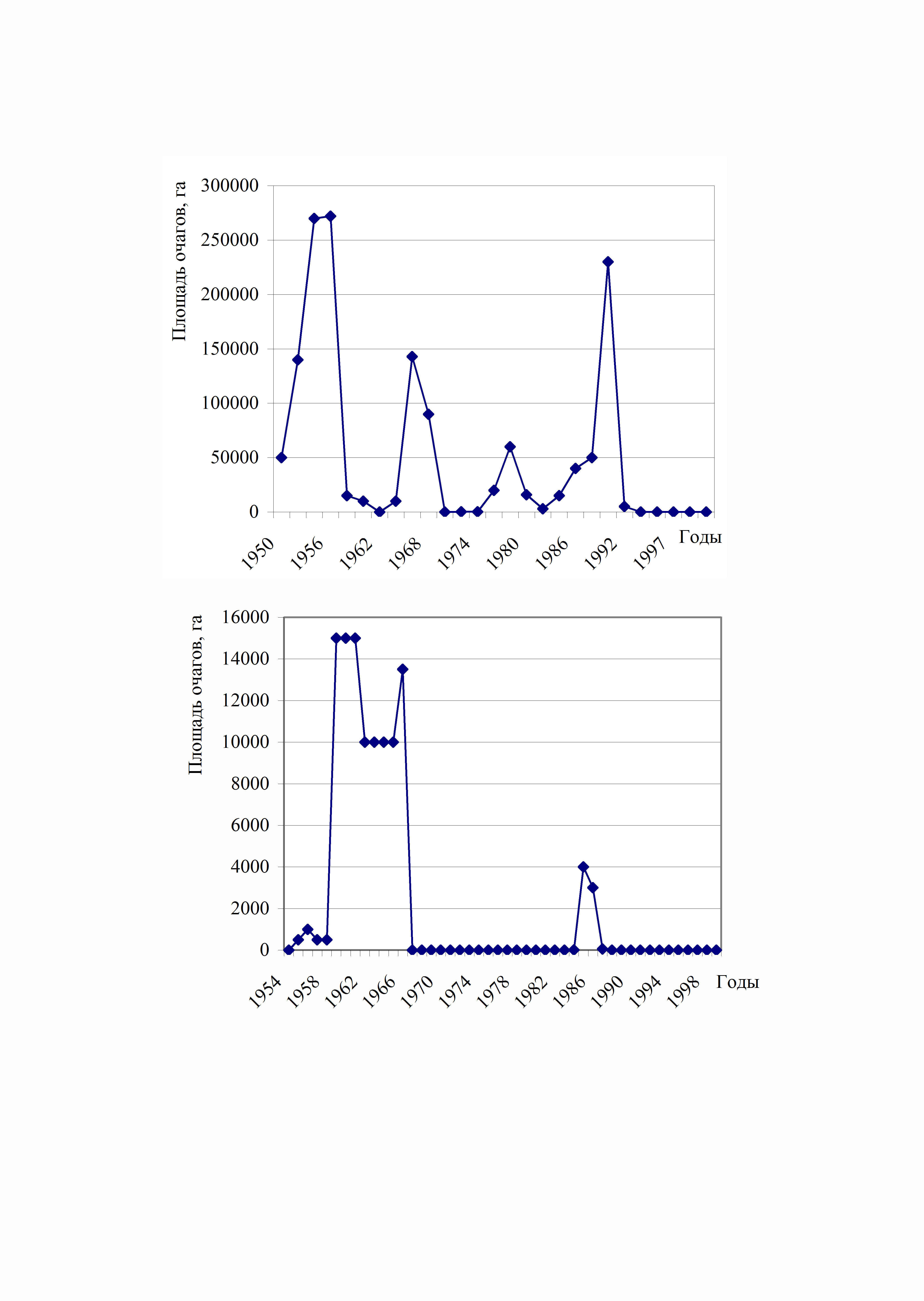
Рис. 4. Динамика вспышек массового размножения непарного
шелкопряда в Челябинской (верхний) и
Свердловской (нижний) областях
Самой крупной по площади и продолжительной была вспышка массового размножения в 1953- 1958 г.г. Это обусловлено синхронизацией вспышки с особенно интенсивными и продолжительными весенне-летними засухами. Вследствие продолжительного воздействия фактора абиотического стресса на древостои наблюдался и частичный отпад березняков (Распопов, 1961, 1970, 1973; Колтунов, 1993,1996; Колтунов и др., 1992, 1998). Второй по интенсивности была вспышка массового размножения в 1984- 1992 г.г. Вспышка была также синхронизирована с засухами (Koltunov, Andreeva, 1999). Более подробно взаимосвязь вспышек массового размножения с засухами рассмотрена в главе 5. Периодичность возникновения очагов массового размножения непарного шелкопряда в лесах Челябинской области составила 9-11 лет (Колтунов и др., 1992,1998; Колтунов, 1993). Предпочитаемые породы: береза, осина.
В лесах Свердловской области первая вспышка массового размножения непарного шелкопряда сформировалась в 1954 г. и продолжалась до 1970 г.г. (рис. 4). Следующая вспышка возникла только в 1986 г и затухла в 1990 г. Популяция непарного шелкопряда обитает только в южной и юго-западной части области (Колтунов, 1993). Это обусловлено более благоприятными климатическими условиями в этой зоне. Периодичность составляет, в среднем, 20 лет. Обращает внимание, также то, что общие площади очагов несопоставимы с очагами в Челябинской области, так как многократно ниже.
В целом такая же асинхронность наблюдается и у группы зауральских очагов массового размножения непарного шелкопряда (рис. 5). Так, если в 1990 г. крупная вспышка массового размножения непарного шелкопряда в Челябинской обл. уже затухает, в лесах Тюменской области она только к этому периоду начинает развиваться (рис. 5). Такая же закономерность наблюдается и в лесах Курганской области, только здесь эруптивная фаза особенно хорошо выражена.
В начале 2007 г. в лесах Курганской обл. выявлено 21.275 га очагов массового размножения непарного шелкопряда, на конец года - 64.382 га; в лесах Свердловской обл. на начало года выявлено 115 га очагов, на конец - 223 га; в лесах Тюменской обл. на начало года обнаружено19105 га очагов непарного шелкопряда, на конец года - 94537 га; в лесах Челябинской обл. на начало 2007 г. выявлено 15404 га очагов, на конец года - 23196 га (Рослесхоз, 2007).
Для изучения факторов асинхронности нами проводилось изучение основных абиотических факторов в фазе депрессии, начала развития вспышки, в эруптивной фазе и фазе затухания на примере очагов массового размножения непарного шелкопряда в Челябинской области (Колтунов и др., 1998; Колтунов, 2006). Было убедительно показано, что характер развития вспышки детерминируется типом и последовательностью сочетания абиотических факторов, которые заметно различаются не только в разных областях, но и в условиях соседних метеостанций одной области.
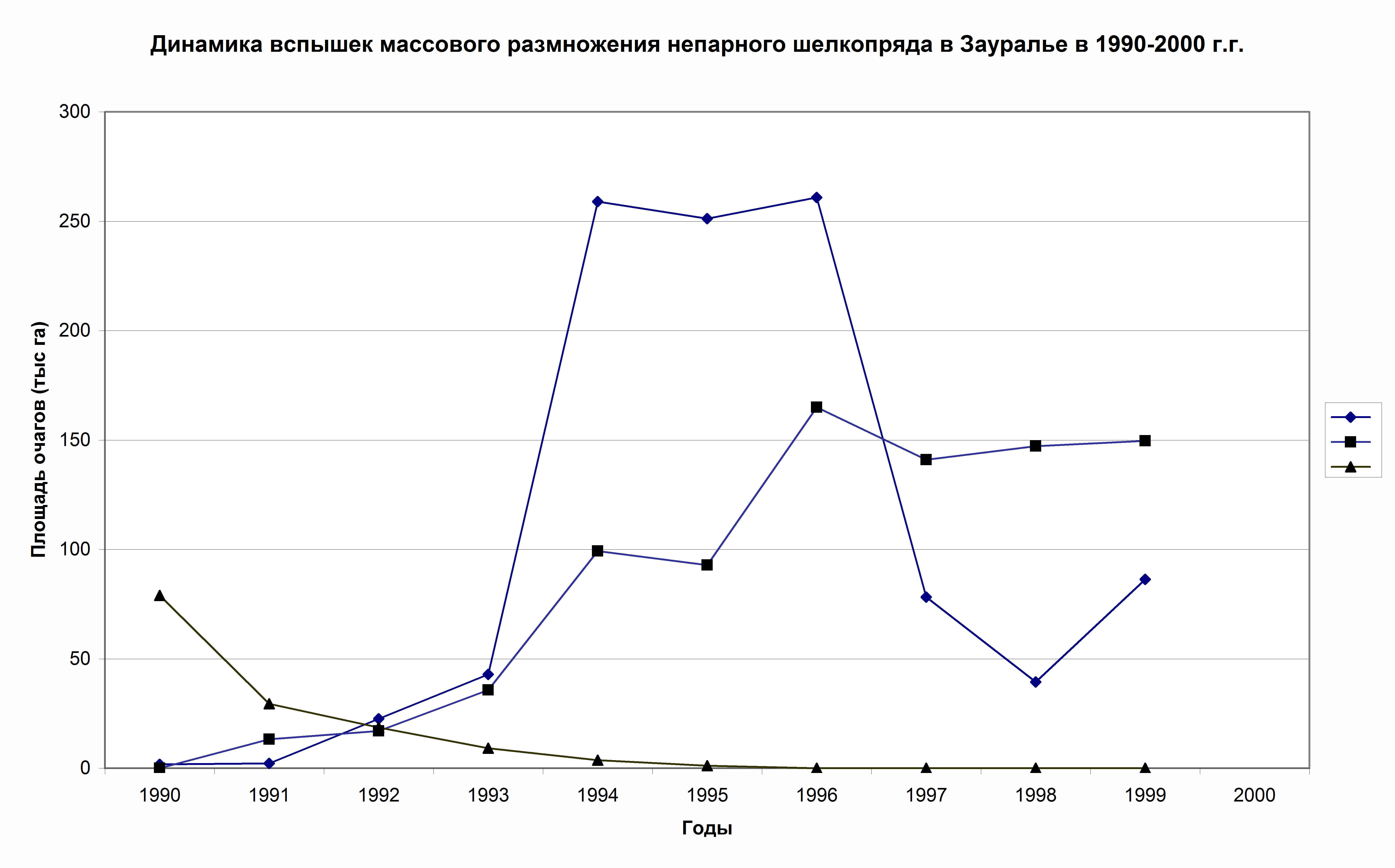
Рис. 5. Особенности популяционной динамики зауральской (▲), башкирской (■) и тюменской (♦) географических популяций непарного шелкопряда с 1990 по 2000 г.г.(Колтунов, 2006)
Таким образом, локальная пространственная структура очагов детерминируется особенностями локального проявления климатических условий (весенне-летних засух и суховеев, уровнем их интенсивности, продолжительности), которые всегда отличаются даже в условиях двух рядом находящихся областей. Чем более интенсивные и продолжительные засухи, тем выше потенциал реализации вспышки и площади очагов и меньше различия в реализации вспышек в рядом расположенных регионах. Достаточно убедительным примером служит вспышка массового размножения непарного шелкопряда в Зауралье в 50-е годы, когда наблюдались особенно интенсивные, стабильные и продолжительные засухи (Колтунов, 1993, 1996, 2006; Колтунов и др., 1998). Более подробно эта проблема обсуждается в данной монографии в главе 5.
Азиатская форма непарного шелкопряда отсутствуют не только в европейской части Российской Федерации, но и в Европе и странах Северной Америки. В странах Тихоокеанского региона этот вид является карантинным. В этих странах включение непарного шелкопряда в список карантинных видов абсолютно обосновано, так как вид либо отсутствует на территории стран, либо имеет ограниченное распространение.
В 1992 году азиатская форма непарного шелкопряда была внесена в «Перечень вредителей растений, возбудителей болезней растений, растений (сорняков), имеющих карантинное значение для Российской Федерации» как объект внешнего карантина. В настоящее время предпринимаются попытки наложения внутреннего карантина в Приморском, Хабаровском краях, а также Республике Саха и некоторых субъектах Приволжского округа в связи с действием в этих регионах очагов непарного шелкопряда. По нашему мнению, которое основано на результатах многолетних исследований экологии непарного шелкопряда (Колтунов, 1993, 1996, 2006; Колтунов и др., 1998) внесение этой формы в список карантинных объектов было совершенно необоснованно. Для специалистов, которые хотели бы получить более полную информацию по этому вопросу, отсылаем к нашей публикации (Колтунов Е.В., Пономарев В.И., Федоренко С.И. О введении карантина против азиатской расы непарного шелкопряда // Лесное хозяйство, №4, с. 43-46, 2001), в которой этот вопрос проанализирован очень детально. Также в статье приведены, по нашему мнению, достаточно убедительные аргументы об ошибочности этого решения. Сходной позиции придерживается в настоящее время и Рослесхоз (Рослесхоз, 2007). К сожалению, необходимо отметить, что внесение этой формы непарного шелкопряда в объекты карантина наносит довольно заметный экономический ущерб России, так как требуется проведение достаточно дорогостоящих мероприятий по карантину.
Рекомендации по защите лесов от непарного шелкопряда
Вряд ли приходится сомневаться, что специалистам по защите леса, интересующимся научной литературой по этому объекту, хорошо известно наше мнение о сложившейся странной ситуации с борьбой против очагов непарного шелкопряда в березовых лесах (Колтунов, 1993, 1996; 2006; Колтунов, Федоренко, 1998; Колтунов и др., 2006). Как известно, береза имеет исключительно высокий уровень энтомотолерантности в благоприятных лесорастительных условиях. Поэтому при однократной полной дефолиации обычно процент усыхания не превышает 2-5% (Колтунов, 1993, 1996, 2006; Колтунов и др., 1998). Особенно это касается устойчивости к дефолиации к насекомым-фитофагам весенне-летней группы. Более заметное локальное усыхание нам удалось обнаружить только в березняках, значительно пораженных бактериальной водянкой после полной дефолиации крон березы непарным шелкопрядом. Повышенный уровень отпада после однократной полной дефолиации березы наблюдался, также, локально, и в результате кислородного голодания корневой системы березы, подвергнутой полной дефолиации на микропонижениях рельефа, вследствие обильных осадков во второй половине лета на почвах, имеющих глиняные линзы в березовых лесах Зауралья (Колтунов, 1993, 1996). Но это имело очень локальный характер. В основном же, подавляющий процент березовых лесов в течение более, чем 20 лет наблюдений очень благополучно перенес несколько полных дефолиаций крон и имеет в настоящее время достаточно высокую полноту (рис. 6).

Рис. 6. Участок березового леса в Каменск-Уральском р-не, перенесший сильную дефолиацию непарным шелкопрядом в 1966 г., 1986 г., летне-осенним комплексом фитофагов в 2000 г. и средний уровень дефолиации в 2006 г., а, также, слабую дефолиацию в 2007-2009 г.г.
Вместе с тем, локально, отпад насаждений, в результате повреждения непарным шелкопрядом в 2007 году, отмечен на 53 га в лесах Курганской и Челябинской областей, где насаждения были предварительно ослаблены от воздействия неблагоприятных погодных условий. Но общая площадь очагов составляла 64 тыс. га (Рослесхоз, 2007). В Приморском и Хабаровском краях за весь период наблюдений (начиная с 50-х годов прошлого века) случаев гибели насаждений не отмечено.
Поэтому массовые обработки очагов против непарного шелкопряда в березовых лесах, которые проводятся и в настоящее время, совершенно экологически и научно необоснованны. В связи с этим мы можем рекомендовать лишь отказ от массовых обработок березняков в очагах массового размножения непарного шелкопряда на территории Уральского Федерального округа.
Вместе с тем, эта рекомендация совершенно не исключает того, что, в определенных условиях на локальном уровне эти обработки можно проводить, например, при необходимости предотвращения дефолиации березовых лесов вокруг детских лагерей, домов отдыха, санаториев и т.п. объектов исключительно для сохранения участков леса в эстетических и рекреационных целях. То же касается и городских лесопарков и зон массового отдыха. Элементы новой стратегии защиты леса, предложенные нами (Колтунов, 1998, 2006), приведены в отдельном разделе сайта.
Совершенно другая экологическая ситуация складывается в очагах массового размножения непарного шелкопряда в дубовых лесах. И, хотя в научной литературе приводятся достаточно различные сведения об уровне энтомотолерантности дуба к дефолиации, тем не менее, сложная ситуация с ареалом дуба в России, особенно в его восточной части, неправильное ведение лесного хозяйства в дубовых лесах (порослевое хозяйство), интенсивное воздействие антропогенных факторов и исключительно реальная опасность сильного временного ослабления дуба после сильной дефолиации, которая часто сопровождается поражением его инфекционными болезнями, по нашему мнению, делает неактуальными научные дискуссии на тему: необходима ли обработка очагов массового размножения в дубовых лесах. Мы считаем, что, в отличие от березы, в дубовых лесах, при опасности сильной однократной дефолиации, или средней двукратной целесообразно проведение обработок очагов для предотвращения сильного временного ослабления дуба и его частичного усыхания и отпада. Краткость рекомендаций в этой части обусловлена, практически, полным отсутствием дуба в лесах Уральского Федерального округа (за очень небольшим исключением).