Шелкопряд-монашенка (Lymantria monacha L.)
Шелкопряд-монашенка (Lymantria monacha L.)
Повреждаемые породы: Cосна и ель; также может повреждать пихту, лиственницу, кедр. Из лиственных древесных и кустарниковых пород повреждает осину, березу, иву, рябину и другие.
Характер повреждений: Гусеницы, выйдя из яиц, питаются молодой хвоей майских побегов, еще не вышедшей из чехликов. Взрослые гусеницы едят старую хвою, перекусывая хвоинки на различной высоте от основания.
Распространение: Во всей Европе, в Западной и Восточной Сибири, Прибайкалье, Приамурье, Приморье, на юге Курил и Сахалина, в Японии и Китае (по крайней мере, в Восточном). В Западной Сибири и Красноярском крае очаги приурочены к спелым и перестойным лесостепным борам, где они действуют практически одновременно с очагами сосновой пяденицы.
Морфология: Бабочки очень изменчивой окраски, с четырьмя черными извилистыми поперечными полосами и черными крапинами на передних крыльях, могут быть беловатые с четким черным рисунком, полностью дымчатые совсем без рисунка и чисто черные.
Задние крылья серые с мелкими черными пятнами по краям (рис. 1,2). Самка с черными нитевидными усиками, с черными и розовыми полосками на брюшке; размах крыльев 40-55 мм. Самец с перистыми бурыми усиками, размах крыльев 25-35 мм.
Яйца почти шаровидные, величиной с маковое зерно, несколько уплощенные с полюсов. Недавно отложенные яйца светло-розовые, затем темнеют. Молодые гусеницы темно-серые с черной блестящей головой, с шестью парами желтоватых бородавок на каждом сегменте тела, на которых расположены длинные, тонкие волоски.
Взрослые гусеницы до 50 мм, серые с желто-бурой головой, волосистые бородавки окрашены в голубовато-серый цвет. Вдоль спины проходит двойная темная полоса, которая на середине тела расходится, охватывая светлое пятно (рис. 3). Куколка коричневая с пучками седых волосков, без кокона, прикрепляется редкими паутинными нитями между хвоинок или на ветвях и стволе (рис. 4).
Жизненный цикл: Бабочки летают с середины июля до середины августа, по вечерам и ночью. Днем неподвижно сидят на стволах деревьев. Самки откладывают яйца кучками от 25 до 145 штук глубоко в щели и трещины коры. В сосняках Сибири яйцекладки располагаются в комлевой части деревьев до высоты 50 см от поверхности земли. В среднем одна самка откладывает 200-300 яиц.
Зимовка протекает на стадии яйца. В начале-середине мая из яиц выходят молодые гусеницы, которые некоторое время остаются вместе, в скоплении, которое называется "зеркало", и лишь через несколько дней поднимаются в крону. Гусеницы проходят пять возрастов и в первой декаде июля приступают к окукливанию в кроне дерева и на подросте.
Ближе к концу июля из куколок выходят бабочки. Учитывая, что повреждение крон шелкопрядом-монашенкой происходит в первой половине лета, гибель насаждений в очагах вредителя наблюдается крайне редко. Обычно в очагах усыхает подрост, деревья верхнего яруса успешно восстанавливаются даже после интенсивных повреждений.

Рис. 1. Бабочка шелкопряда-монашенки

Рис. 2. Бабочка шелкопряда-монашенки темного фенотипа (фото: В. Гуменюк)

Рис. 3. Гусеница шелкопряда-монашенки (фото: Stanislav Kinelski, Bugwood. org)

Рис. 4. Куколка шелкопряда-монашенки
Динамика вспышек массового размножения шелкопряда-монашенки
в лесах Зауралья. Челябинская обл.
Проблемы популяционной экологии шелкопряда-монашенки исследуются очень давно (Тур, 1893, 1894; Филипьев, 1894; Прозоров, 1949;Распопов,1961; Ханисламов и др., 1962; Черепанов, 1963; Трифонов др., 1991; Соколов, Бахвалов,1992; Бахвалов, Соколов, Солдатов, 1992; Бахвалов и др., 1990; , Ильиных, Бахвалов, Божко,1990; Максимов: 1992; Schonherr,1985, 1989; Sliwa, 1987, 1994; Jensen, 1991, 1996). Но в условиях воздействия антропогенных факторов многие аспекты популяционной динамики остаются недостаточно изученными.
Первые упоминания о вспышках массового размножения шелкопряда-монашенки в Зауралье относится к 1889 г. (Распопов, 1961). Примерно к этому же времени (1894г.) относятся и первые сообщения о появлении этого фитофага в Оренбургской области (Симон, 1895). О вспышках массового размножения шелкопряда-монашенки в Пермской губернии (в состав которой входила и Свердловская область) сообщал и Тур (Тур, 1894). Вспышки массового размножения зауральской популяции шелкопряда-монашенки в Челябинской области наблюдались в 1951-54 годах, 1979 и 1985-1989 годах (Распопов, 1961; Соколов, 1990).
В Предуралье, в лесах Башкирии, в течение последних 40 лет также наблюдались вспышки массового размножения шелкопряда-монашенки: в 1956-1976-1984 и 1988-1991 г.г. (Ханисламов и др., 1962; Трифонов и др., 1991). Шелкопряд-монашенка постоянно обнаруживался в резервациях. Резервациями этого фитофага в Предуралье служат средневозрастные высокополнотные сосняки (Трифонов и др. 1991).
В Зауралье у популяции шелкопряда-монашенки вспышки массового размножения формировались, преимущественно, в спелых древостоях. Лишь в последние годы очаги стали обнаруживаться в культурах сосны обыкновенной, возрастом старше 10 лет (Соколов, 1990). Другие породы хвойных в Предуралье и Зауралье этот фитофаг не повреждает (Ханисламов, 1962; Трифонов и др., 1991). Хотя известно, что этот фитофаг способен повреждать, также, кедр, темнохвойные, а в условиях Дальнего Востока и листву дуба. Интересно отметить, что общая площадь очагов шелкопряда-монашенки в Башкирии, выявленная в 1990 г. составляла всего 3130 га (Трифонов и др., 1991), что было гораздо меньше общей площади очагов этого фитофага в Челябинской области в этом же году. Вероятно это обусловлено более аридными условиями в сосновых борах Челябинской области, по сравнению с Предуральем. Ряд специалистов связывает разную частоту и продолжительность вспышек массового размножения шелкопряда-монашенки с составом древостоя.
По мнению J. Schonherr (1989) в сосновых лесах периодичность вспышек массового размножения составляет 4 года, в еловых лесах - 7 лет (Schonherr,1989). Зоны вспышек массового размножения этого фитофага приурочены в Европе к южным и восточным районам (Schonherr ,1989).
Исследования в сосновых лесах Северного Казахстана показали, что плодовитость самок в культурах сосны была выше чем в естественных сосняках (Бахвалов и др., 1989). В период вспышки 1984-1986 гг. в культурах 35 летнего возраста сильная ежегодная дефолиация не наблюдалась (Бахвалов и др., 1989).
Известно, что сосна обыкновенная обладает относительно высокой устойчивостью к дефолиации насекомыми-фитофагами (Воронцов, 1978; Тальман, Яцентковский, 1938). Причем этот факт отмечается в течение очень длительного периода, превышающего 100 лет (Воронцов, 1978). Анализ воздействия фактора дефолиации крон сосны обыкновенной шелкопрядом-монашенкой в Зауралье показал, что отпада древостоя, как правило, не происходит (Бахвалов и др., 1989, 1992; Соколов, Бахвалов, 1992; Максимов, 1992,1994). Лишь в ряде случаев в сосновых культурах после дефолиации их в течение 3-х лет он составлял не более 5-10%. Причем в составе отпада были усыхающие, угнетенные деревья 3 и 4 классов возраста (Бахвалов и др., 1989; Соколов, Бахвалов, 1992).
Исходя из этого, совершенно очевидно, что уровень энтомотолерантности сосны обыкновенной в Зауралье высок. Поэтому сильного отпада, как правило, не происходит. Повышенный процент отпада обычно связан с различными дополнительными факторами ослабления древостоев (Соколов, Бахвалов, 1992). Но имеются, также, и другие результаты, свидетельствующие о значительном отпаде сосны обыкновенной после дефолиации сосновой пяденицей и сосновым шелкопрядом (Воронцов, 1978). П.М. Распоповым, также, было показано, что сильная дефолиация кроны сосны обыкновенной в условиях лесостепи (Уйский и Варламовский боры) сопровождалась тем, что полное восстановление радиального прироста сосны происходило в течение длительного времени, а древостои восстановились через 4-5 лет.
После вспышки в отдельных древостоях возникли очаги стволовых вредителей (Распопов, 1961). Сходные результаты получены также М.Г. Ханисламовым с сотрудниками (Ханисламов и др., 1962). Сравнение параметров устойчивости сосны к дефолиации этим фитофагом с европейскиими популяциями затруднительно, так как шелкопряд-монашенка распространен в Европе, преимущественно, в еловых лесах (Гловацка, 1989; Sliwa, 1987, 1994; Schonherr, 1972, 1989; Grjpma, 1989; Jensen, 1991, 1996).
Сравнительный анализ устойчивости сосны обыкновенной к дефолиации шелкопрядом-монашенкой из других популяций в России показывает, что разными авторами приводятся заметно различающиеся результаты (Шевырев, 1894; Воронцов: 1978; Авраменко и др., 1981; Знаменский, Белов, 1981). По нашему мнению, это свидетельствует в большей степени не о действительно заметных различиях в уровне энтомотолерантности сосны обыкновенной в разных условиях произрастания, а о том, что в разные вспышки, сопряженные с различным уровнем воздействия фактора абиотического стресса (засухи), устойчивость сосны к дефолиации может различаться.
Известно, что после повреждения хвои сосновой совкой древостой восстанавливаются хуже, при дефолиации другими фитофагами (Воронцов, 1963,1978). Это обусловлено, в первую очередь, способностью сосновой совки к уничтожению почек (Тальман, Яцентковский, 1938).
Так А.И. Воронцовым при обследовании сосны обыкновенной в очагах массового размножения шелкопряда-монашенки в Татарской АССР в 1942 гг. и в Свердловской области в 1953-54 гг. ни усыхания древостоев, ни повреждений их насекомыми-ксилофагами не обнаружено (Воронцов, 1978). Автор предполагает, что это могло быть обусловлено влажной погодой в период затухания вспышки.
Известны также отдельные факты значительного отпада сосны обыкновенной в 90-х годах 19 века после дефолиации шелкопрядом-монашенкой в Пермской губернии (Тур,1894). Следует отметить, что фактических данных по воздействию фактора дефолиации на сосну обыкновенную в Зауралье в период вспышки массового размножения в 50-е годы очень мало (Распопов, 1961). В целом мнения многих специалистов сходятся на том, что усыхание и отпад сосны обыкновенной наблюдается лишь при совпадении нескольких отрицательных факторов одновременно: двух-трехкратной дефолиации, засухи и высокой численности насекомых-ксилофагов (Старк,1932; Прозоров, 1956; Воронцов, 1978).
Таким образом, в целом особенности популяционной экологии шелкопряда-монашенки в Зауралье изучены очень фрагментарно. Единичные публикации относятся, в основном, к 50-60 годам. Тем не менее, тесная взаимосвязь вспышек массового размножения этого фитофага в Зауралье с весенне-летними засухами установлена достаточно убедительно (Распопов, 1961).
Взаимосвязь интенсивности засух с интенсивностью развития очагов, масштабами вспышек и лесорастительными условиями, также была установлена (Распопов, 1961), что имеет важное значение для понимания ключевых факторов популяционной динамики этого фитофага в условиях Зауралья. Но факт синхронизации появления очагов с весенне-летними засуха-

Рис. 5. Динамика вспышек массового размножения шелкопряда-монашенки в сосновых лесах Челябинской обл. (1951-2008 г.г.)
ми совершенно недостаточен для реализации механизма вспышек, предложенного нами ранее для непарного шелкопряда (Колтунов, 2006). Как показали результаты, полученные нами ранее, для успешной реализации вспышки необходима синхронизация засух с периодом, предшествующим подъему численности популяции, продромальной и эруптивной фазам, а, также, определенный тип весенне-летних засух, их характер, интенсивность и продолжительность.
Анализ особенностей периодичности вспышек массового размножения шелкопряда-монашенки в Зауралье за последние 53 года убедительно показывает, что она не была стабильной, а заметно различалась. Так, в сосновых лесах Челябинской обл. первая зарегистрированная вспышка массового размножения этого фитофага возникла в 1951 г и продолжалась до 1956 г (рис. 5).
Затем в 1958 г. она возникла вновь и продолжалась до 1961 г. При этом по характеру вспышка массового размножения была очень интенсивной, а площадь очагов была высокой (32 тыс. га). Столь узкий период депрессии, вероятно, обусловлен необычно сильными и продолжительными весенне-летними засухами, по мнению многих авторов.
Как показали результаты (рис. 5) следующая вспышка массового размножения наблюдалась в сосновых лесах Челябинской обл. лишь в 1976 г., после фазы депрессии, продолжавшейся в течение 15 лет. Она затухла в 1983-1984 г.г. Площадь ее была незначительной (6 тыс. га). Следующая вспышка массового размножения возникла в 1987-1988 г.г. и затухла в 1997-1998 г.г. Ее площадь была очень значительна (33 тыс. га) и сравнима с площадью очагов массового размножения в 50-х и 60-х годах (рис. 5). Последняя вспышка массового размножения вновь возникла в 2003 г и продолжается до 2009 г. Ее площадь была очень незначительна.
Анализ гидротермических условий в этот период убедительно показывает, что весенне-летние засухи наблюдались в 1967-1968 г.г.. Затем, в 1969-1972 г.г. они сменились периодом влажных лет, с очень высокими гидротермическими коэффициентами мая-июня. Здесь целесообразно проанализировать: отличались ли засухи 1967-1968 г.г. по характеру, интенсивности. Продолжительности с предыдущим циклом весенне-летних засух, синхронизированных со вспышкой массового размножения 1951-1956 г.г.
Заслуживает внимания, что столь длительный интервал между вспышками, наблюдавшийся в Зауралье, был характерен не только для зауральской популяции шелкопряда-монашенки, но наблюдался и в восточно-европейских популяциях этого фитофага. Так в Польше после 40-х годов очередная вспышка массового размножения возникла только в 1977 году (Schonherr, 1985).
Одной из причин мы считаем проведение широкомасштабных обработок очагов инсектицидами в биогеоценозах Зауралья в этот период (Распопов, 1961). Как известно, с 1953 по 1956 г. в сосновых лесах Челябинской области проводились массовые авиахимические обработки против очагов шелкопряда-монашенки. За этот период было обработано 50,7 тыс. га лесов (Распопов, 1961). В Восточной Европе также очень широко применялась обработка инсектицидами очагов шелкопряда-монашенки. Например, в Польше с 1978 по 1982 г. было обработано более 2,5 млн. га лесов (Schonherr, 1985). После длительного перерыва очаги массового размножения шелкопряда-монашенки в Польше возникли на огромной территории (2 млн. га (Schonherr, 1985).
Исследования показали, что в период депрессии (1962-1973г.) во многих районах Челябинской области яйцекладки шелкопряда-монашенки не обнаруживались. Фаза депрессии численности шелкопряда-монашенки была тесно синхронизирована с фазой максимума гидротермических коэффициентов (периодом повышенной влажности) и кратковременными засухами в 1963-1965 г.г., которые часто прерывались месяцами с высокими ГТК. Незначительный подъём численности зарегистрирован в 1973 году в Курганской области (Долматовское, Курганское лесничества и в Челябинской области: в сосновых лесах Увельского района. Позже он наблюдался в лесах Ларинского лесничества Чебаркульского лесхоза и в Кыштымском бору Кыштымского лесхоза (рис. 6). В период лёта имаго в 1973 году в этих сосновых борах было обнаружено 0,01-0,03 бабочек на 1 дерево.
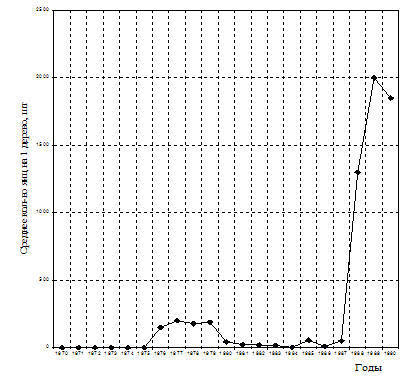
Рис. 6. Особенности популяционной динамики шелкопряда-монашенки в сосновых лесах с наибольшей площадью очагов (1970-1990 г. г.) Челябинская обл. Чебаркульский ОПЛК.
Несмотря на то, что возрастание численности популяции в разных районах происходило неравномерно, тем не менее, уже с 1971 года начинается рост численности в популяции в целом. Так, если в 1970 году кладки вообще не обнаруживались, то в 1971 году среднее количество яиц (по данным осенних учетов) на 1 дерево составило: 0,35; в 1972-2,5, в 1973 г. они вновь не обнаруживались на учётных площадях, а в 1974-1,4 (рис. 6, 7). Еще через год в отдельных кварталах Варламовского лесничества Чебаркульского ОПЛХ обнаруживалось в среднем 7,5 яиц на дерево, а в лесах Увельского лесхоза - 9,8 шт. В 1976-77 гг. средняя численность популяции резко возросла до 144 яиц на дерево. Происходит интенсивное формирование очагов (рис. 6, 7).

Рис. 7. Особенности популяционной динамики шелкопряда-монашенки в длительно действующих очагах. Челябинская обл. Анненский лесхоз. 1970-1991 г. г.
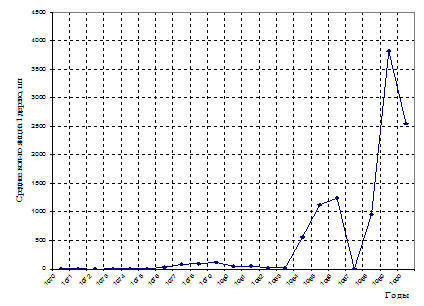
Рис. 8. Особенности популяционной динамики шелкопряда-монашенки в длительно действующих очагах. Челябинская обл. Еткульский лесхоз. 1970-1991 г. г.
В 1977 г. первые очаги массового размножения шелкопряда-монашенки возникли в сосновых лесах Чебаркульского ОПЛК (на площади 400 га) и Еткульском лесхозе (на площади 100 га) (рис. 8). На следующий год новые очаги сформировались в сосняках Анненского лесхоза, а площадь очагов, возникших в 1977 г. быстро возросла.
Общая площадь очагов в этот период составляла 5.826 га (рис. 6-8). Как следует из приведенных данных общая площадь очагов шелкопряда-монашенки в период вспышки 1977-1983 г.г. была очень незначительной. Очаги, возникшие в этот период были, в основном, мелкими по площади, за исключением очагов в лесах Чебаркульского ОПЛК, где встречались и крупные очаги (рис. 6-8,11). Но очаги в Чебаркульском ОПЛТК были в период этой вспышки не только наиболее крупными по площади, но и одними из самых длительных по продолжительности.
Общая продолжительность вспышки в сосняках Чебаркульском ОПЛК составляла 6 лет (рис. 11). В локальных очагах степень дефолиации была либо слабой (до 30%), либо умеренной (до 50%). Лишь на 20% площади очагов ожидалась сильная дефолиация кроны.
Следует, также, обратить внимание на то, что участки, где был обнаружен ранний подъем численности популяции шелкопряда-монашенки в 1972-1973 г.г., пространственно не совпадали с древостоями, где формировались первые очаги массового размножения в 1977-1978 г.г. .
Обращает внимание и то, что, если более половины насаждений, где зарегистрировано начало подъема численности зауральской популяции шелкопряда-монашенки расположены в северной части Челябинской области, в лесах Курганской области, граничащих с территорией Челябинской области, в северной подзоне лесостепи, а один участок - в лесной, то первые очаги формировались, преимущественно, ближе к южной границе лесостепной зоны, а также - в степной зоне (Колтунов, 1996).
Как показали результаты вспышка 1977 г. была слабой, площади дефолиированных древостоев были незначительны (рис. 6-8, 11) за исключением лесов на территории Чебаркульского ОПЛК (4.055 га).
Мы предполагаем, что обнаруженные нами особенности характера пространственной структуры популяции в период подъема численности и формирования первичных очагов могут быть обусловлены особенностями локального проявления засух, как ключевого стрессового фактора. Так ранее для зауральской популяции непарного шелкопряда нами установлено, что формирование совершенно определенной пространственной структуры первичных очагов детерминируется дифференцированной реакцией на стресс (возникновение засухи) древостоями, растущими в разных почвенных условиях (Колтунов, 1993).
Кроме того, ранее П.М. Распоповым было показано, что в условиях слабых засух очаги массового размножения этого фитофага образуются, в основном, в степной и в меньшей степени, лесостепной зонах, а при сильных засухах достигают лесостепи, горно-лесной зоны (Распопов, 1961, 1970, 1973). Результаты, полученные нами, хорошо согласуются с этими данными.
В 1979 г. самые поздние очаги сформировались в сосновых борах Кизильской ЛСС, на общей площади 442 га и в Багарякском лесхозе, площадью 629 га. В 1980 году сформировался очень мелкий очаг в Брединском р-не, площадью 32 га (рис. 8). Еще позже, в 1982 году его площадь увеличивается до 816 га, тогда как самый крупный - Чебаркульский очаг году полностью затух.
В целом анализ пространственно-временной структуры вспышки массового размножения шелкопряда-монашенки 1977-1980 гг. позволил установить, что первые очаги, как правило, формировались в центре ареала зауральской популяции шелкопряда-монашенки. В течение всего периода вспышки наибольшее количество очагов и наибольшая их площадь была также в центре ареала популяции (Колтунов, 1996). Формирование самых поздних очагов происходило не только ближе к северной границе ареала популяции, как можно было предполагать, но с одинаковой интенсивностью происходило и на юге ареала, в степной зоне. Общая площадь очагов в степной зоне была наименьшей (136 га). Очаги с максимальной площадью возникли в Чебаркульском ОПЛХ и Уйском лесхозе (общая площадь - 5250 га) (рис. 7,8, 11).
Как показали результаты анализа топографии очагов характерной особенностью данной вспышки является то, что даже в 1982-83 гг. полного затухания очагов не произошло, так в 1984 г. вновь стал наблюдаться площади очагов. В 1988 г. общая площадь очагов вновь резко возка и достигла 16047 га, в 1989 - 26624 га, в 1990 - 32223 га. Следовательно, вспышка 1977 года фактически трансформировалась без полного затухания в следующую, гораздо более крупную по площади вспышку (рис. 11).
Сравнительный анализ характера динамики численности шелкопряда-монашенки в разных частях ареала показывает, что непосредственный переход в фазу новой вспышки свойственен для популяции в целом. По этим районам обнаруживается очень широкий диапазон характера популяционной динамики: от непосредственного перехода к подъёму численности, без фазы полного затухания до довольно длительной, многолетней фазы затухания, когда очаги не обнаруживаются в сосняках в течение очень многих лет, до вспышки 1989-1990 гг. (рис. 7,8). При тесной корреляции с особенностями локальных очагов наблюдается совершенно различный характер динамики заселенности древостоев яицами шелкопряда-монашенки.
Так в длительно действующих, где вспышка продолжается 10-15 лет (Еткульский очаг), (рис. 8) показатель средней заселенности древостоев яйцами, тем не менее, испытывает очень резкие градации (рис. 7). Так в 1982-1985 годах среднее количество яиц резко сокращается до 30-100 на 1 дерево, затем в 1986-1987 возрастает - до 2067 шт. на 1 дерево, и в 1987-88 вновь снижается, до 9, а к 1990 г. резко увеличивается до 2438,0 шт. на дерево (рис. 7).
Совершенно очевидно, что средние показатели плотности популяции шелкопряда-монашенки отражают особенности локального пространственного распределения популяции этого фитофага. Неравномерность заселения сохранялась всегда, даже в эруптивной фазе. В Брединском очаге, например, если в 1982 г. средняя плотность популяции составила 494,1 шт. на 1 дерево, то в 1983 г. очаги затухли и до 1989 г. плотность популяции колебалась от 9,2 до 29,8 яиц на дерево. К вспышке 1989-1990 г. она возросла до 1565 яиц, а общая площадь очагов составила 105 га.
Как показывают результаты, в низкорезистентных сосняках, где периодически возникают вспышки массового размножения, шелкопряд-монашенка обнаруживается постоянно, хотя плотность его популяции варьирует в очень широких пределах. В то же время в горно-лесной зоне, где вспышки возникают очень редко, он не всегда обнаруживается в межвспышечный период. Это не является свидетельством полного отсутствия вида в этих экотопах. Наиболее часто численность популяции столь мала, что она может обнаруживаться только при осмотре очень большого количества деревьев, что связано со значительной трудоёмкостью при проведении обследований.

Рис. 9. Особенности вспышки массового размножения шелкопряда-монашенки в лесах Челябинской обл. в быстротекущих очагах. Слева Верхнеуральский лесхоз, справа – Пластовский лесзоз.
Как показали результаты, вспышка массового размножения 1988-1991 г.г. в отдельных лесхозах возникла сразу же, после затухания предыдущей. Но такой факт наблюдался не во всех лесхозах. Наоборот, во многих из них эта вспышка возникла после достаточно длительного периода фазы депрессии.

Рис. 10. Особенности вспышек массового размножения шелкопряда-монашенки в лесах Челябинской обл. в длительно действующих очагах.
Кроме того, описанным вспышкам всегда предшествовали гидротермические условия, инициирующие абиотический стресс у древесных растений, изменение биохимического состава кормового субстрата, которые быстро отражались на росте выживаемости и экологической плотности популяций шелкопряда-монашенки.

Рис. 11.Особенности вспышек массового размножения шелкопряда-монашенки в лесах Челябинской обл. с максимальной площадью очагов. Чебаркульский ОПЛТК.
Обращает, также, внимание то, что популяция шелкопряда-монашенки очень быстро откликается на изменение гидротермических условий в экотопах изменением экологической плотности в ту, или другую сторону. Исходя из этого можно предполагать, что зауральская популяция шелкопряда-монашенки в климатических условиях, характерных для степной и лесостепной зон, адаптировалась к сильному отклику на изменение гидротермических условий. Поэтому крупномасштабные вспышки возникают обычно только в условиях, когда перед подъемом численности популяции происходит возникновение весенне-летних засух. Особенно майских. В продромальной фазе происходит возникновение часто полных и очень интенсивных весенне-летних засух, редко прерываемых периодом с нормальными, или повышенными ГТК. В эруптивной же фазе такие гидротермические условия складываются почти всегда. Чем более интенсивные и полные весенне-летние засухи (с ГТК 0,2-0,3), тем более интенсивная вспышка массового размножения и большие площади очагов.
Столь быстрый отклик популяции шелкопряда-монашенки на изменение гидротермических условий наводит на предположение о том, что, несмотря на то, что хвоя у сосны многолетняя, ее биохимический состав (в весенне-летний период) все же быстро изменяется, в зависимости от складывающихся в экотопах гидротермических условий. Следовательно, для возникновения вспышки массового размножения у шелкопряда-монашенки совершенно необходимо достаточно интенсивное воздействие фактора абиотического стресса, и в первую очередь, в весенний период (май). Только отклик части ценопопуляции сосны на фактор абиотического стресса сопровождается быстрыми изменениями в биохимическом составе хвои и временным снижением энтоморезистентности древостоев. По крайней мере, именно это установлено нами при изучении взаимоотношений в системе: «береза-непарный шелкопряд» (Koltunov, Andreeva, 1999; Колтунов, 2006).
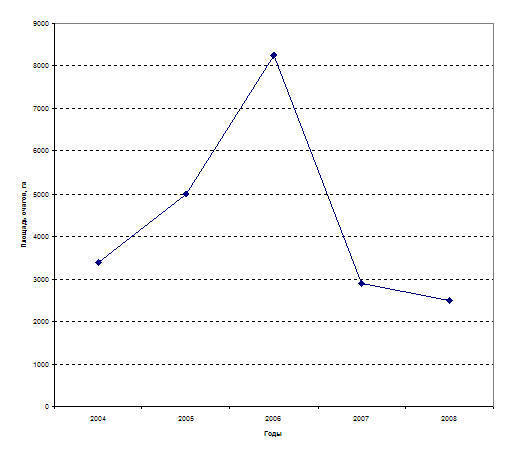
Рис. 12. Особенности вспышки массового размножения шелкопряда-монашенки в сосновых лесах Челябинской обл. в 2003-2008 г.г.
Последняя вспышка массового размножения шелкопряда-монашенки в сосновых лесах Челябинской обл. возникла в 2002 г. и продолжается по настоящее время (2009 г.) (рис. 12).
Особенности популяционной динамики шелкопряда-монашенки
в сосновых лесах Свердловской обл.
Как показал анализ особенностей популяционной динамики популяции шелкопряда-монашенки в Свердловской области за последние 53 года (1955-2008 г. г.), в сосновых лесах области зарегистрированы четыре вспышки массового размножения. Первая вспышка возникла в 1952 г. и затухла в 1956 г. (рис. 13), следующая возникла в 1972-1980 г.г., затем в 1984-1991 г. (рис. 14) и последняя возникла в 2001 г. и в 2008 постепенно затухает, но не полностью.
Фактически на незначительной площади она продолжается и в настоящее время. Следовательно, периодичность возникновения вспышек массового размножения зауральской популяции шелкопряда-монашенки в лесах Свердловской области за последние 45 лет была нестабильной, так как период между первыми двумя вспышками составлял, в среднем, 22 года, тогда как между следующими вспышками - 10-11 лет (рис. 13, 14). Важно отметить, что такая же нестабильная периодичность, как уже отмечалось выше, наблюдалась и у популяций этого фитофага в Восточной и Западной Европе (Grjipma, 1989; Schonherr, 1985, 1989; SSliwa,1987; Jensen, 1991,1996).
Сравнительный анализ продолжительности вспышек за последние 45 лет показал, что она заметно различалась. Так первая вспышка продолжалась в течение 4-х лет, вторая - в течение 9 лет и третья - также в течение 9 лет (рис. 13, 14). Причем характерной особенностью второй и третьей вспышек было то, что как такового хорошо выраженного периода депрессии между вспышками, по существу, не наблюдалось. Поэтому вспышка массового размножения 1972-1980 г.г. г.г. быстро сменилась новой вспышкой в 1983 году (рис. 14). Но даже в межвспышечный период 1982 г.г. в целом для популяции шелкопряда-монашенки в Свердловской области было характерно состояние повышенной численности, а в отдельных сосняках регистрировались локальные очаги на незначительной площади.
Анализ закономерностей развития вспышек массового размножения зауральской популяции шелкопряда-монашенки в Свердловской области показал, что в период первой вспышки 1952-1955 г.г. первые, незначительные по площади очаги возникли в 1951 г. в сосновых лесах Тугулымского, Талицкого и Каменск-

Рис. 13. Особенности вспышки массового размножения шелкопряда-монашенки в лесах Свердловской обл. в 1952-1956 г.г. ■-Талицкий, ▲- Тугулымский, ♦ - Свердловский лесхозы
Уральского лесхозов. Интенсивность подъема численности популяции была столь высока, что уже на следующий (1952 г.) площадь очагов в этих лесхозах многократно возросла и ла: в Тугулымском - 14.427 га, в Талицком - 20.800 га, Каменск-Уральском - 1017 га (рис. 13). Одновременно наблюдается возникшие и группы новых очагов. Очень крупный новый очаг был обнаружен в сосновых лесах Свердловского совхоза, на площади 10.800 га. Зачительный очаг сформировался также в сосняках Сысертского лесхоза (3.000 га) (рис. 13). Следует отметить, что, вероятно, вследствие особенно интенсивного возникновения засухи одновременно на значительной части юга и юго-востока Свердловской области, в первый год возникновения очагов полностью отсутствует обычно характерная тенденция для шелкопряда-монашенки в Свердловской области к более раннему формированию очагов в южной части области (рис. 13).
Все основные очаги сформировались одновременно как в южной и области (Сысертский и Каменск-Уральский районы), так и в Талицком и Тугулымском районах, расположенных северо-восточнее (Колтунов, 1998). При этом площадь очагов была значительно больше именно в двух последних лесхозах (рис. 13).
Безусловно, это связано не только с климатическими особенностями этих районов, особенностями наступления засухи в разных районах, но и особенностями лесорастительных условий.
Так характерно, что в сосновых лесах Сысертского лесхоза, который расположен в подзоне южной тайги, лесорастительные условия в целом менее благоприятны для формирования крупных по площади очагов ной дефолиацией. Поэтому в период всех вспышек массового размножения шелкопряда-монашенки за последние 45 лет в подзоне южной тайги крупных очагов никогда не возникало (Колтунов, 1998).
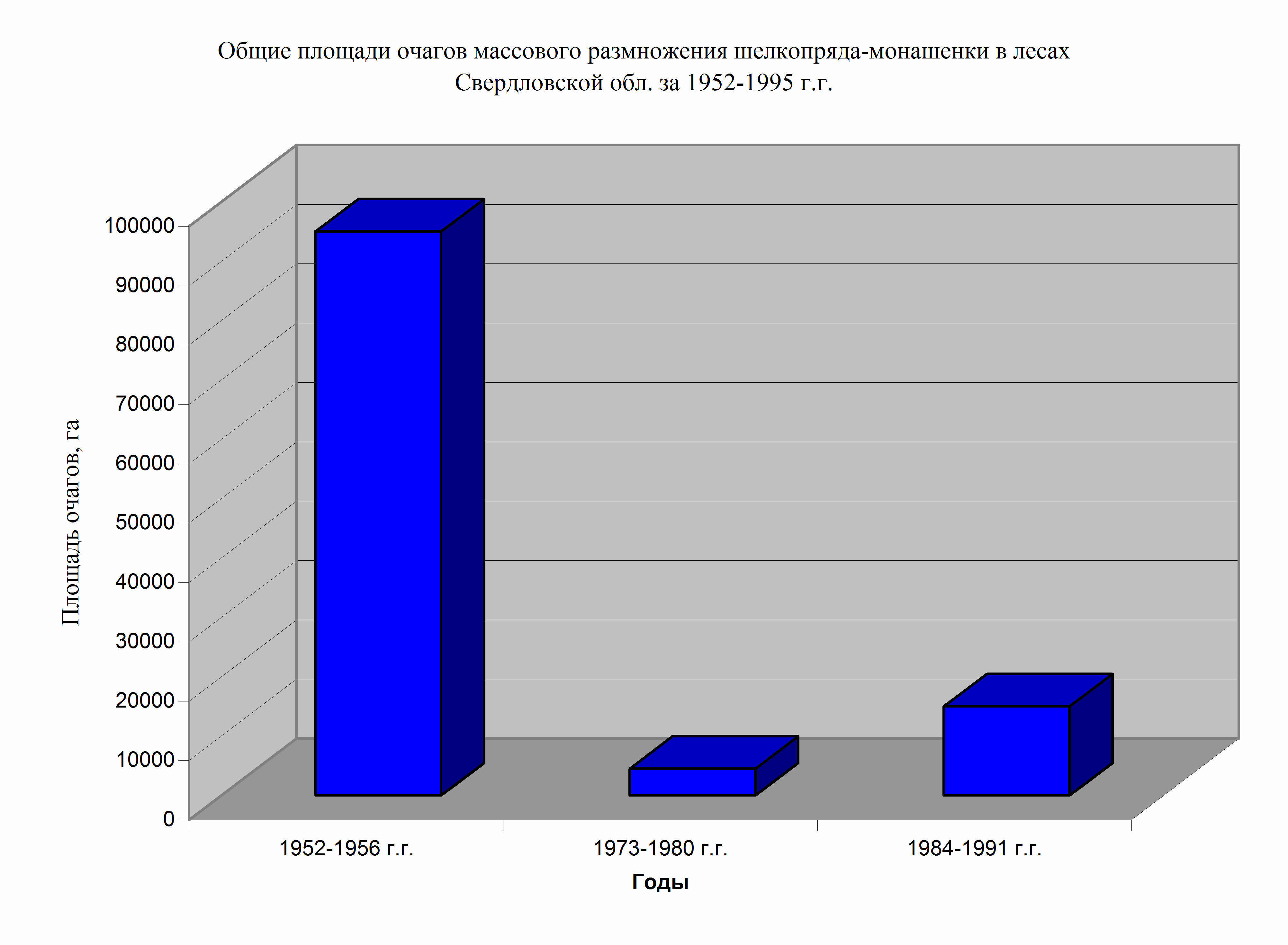
Рис. 14. Общие площади очагов массового размножения шелкопряда-монашенки в лесах Свердловской обл. в 1952-1995 г.г.
Еще через год (в 1953 г.) новые очаги возникли в лесах Березовскогога), Сухоложского (4.400 га), Верхне-Пышминского (284 га) Уралмашевского лесхозов (1211 га). В сосновых лесах с наиболее значительной площадью очагов, возникших ранее, их общая площадь не возрастала (рис. 73). Кроме того новый очаг шелкопряда-монашенки был обнаружен в лесах Камышловского лесхоза. Его общая площадь составляла 4.079 га. Обращает внимание, что почти все эти очаги, возникшие наиболее поздно, сформировались в подзоне южной тайги, что очень несвойственно данной популяции шелкопряда-монашенки при менее интенсивных вспышках массового размножения.
Как показали результаты, другой характерной особенностью динамики пространственной структуры очагов является очень заметная тенденция к продвижению очагов к северной границе ареала популяции. Так, в 1954 году новый очаг, площадью 2239 га, возник в сосновых лесах Алапаевского и Режевского лесхозов (540 га). Особенно важно отметить, что никогда больше в течение последних 53 лет на такой северной широте не наблюдалось формирования очагов массового размножения у шелкопряда-монашенки в Свердловской области. Кроме того на третий год развития вспышки массового размножения (1954 г.) наблюдался значительный рост общей площади очагов в лесах Талицкого района Свердловской обл. Суммарная площадь очагов в этом районе возросла до 40.750 га (рис. 13). Многие из очагов, расположенных в Талицком районе, имели значительный уровень дефолиации крон. Наблюдался также и дальнейший рост общей площади очагов в Тугулымском лесхозе (до 19.800 га) (рис. 13). Все другие очаги, возникшие ранее, не имели тенденции к возрастанию, (рис. 13). В течение 1955 года регистрируется фаза кризиса и затухание вспышки.
Площадь очагов в большинстве лесов быстро снижается, особенно интенсивно этот процесс наблюдается в сосновых лесах подзоны южной тайги, но развивается также и в лесах подзоны северной степи и непосредственно примыкающих к ней. Площади очагов в этот период быстро снижаются до 300-400 га при слабой дефолиации крон древостоев. В 1956 г. происходит полное затухание очагов массового размножения шелкопряда-монашенки в лесах Свердловской области. Наблюдается хорошо выраженная и быстро протекающая фаза кризиса вспышки (рис. 13). В 1957 г. очаги полностью затухли и плотность популяции этого фитофага быстро падает до 2-9 яиц на 1 дерево. В последующие 2 года происходит ее дальнейшее снижение. Как показали результаты в период вспышки массового размножения 1956 г.г. очаги возникали в спелых и перестойных сосновых лесах, в суходольных типах леса. Заметного отпада древостоев после дефолиаации не наблюдалось.
Новая вспышка массового размножения у шелкопряда-монашенки в Свердловской области возникла в 1972 г. в сосновых лесах Камышловского лесхоза. Эта вспышка была очень незначительной по площади - (300 га) и интенсивности протекания. В течение последующих 5 лет, до 1979 г. новых очагов не обнаруживалось. В Камышловском лесхозе площадь очагов в течение 5 лет не изменялась. Лишь в 1979 г. наблюдается формирование новых очагов в лесах Каменск-Уральсского лесхоза, на общей площади 3.500 га. Эта группа очагов продолжала обнаруживаться до 1984 года, но их площадь упала до 500 га (рис. 15). После этого популяция вновь перешла в фазу подъема численности.
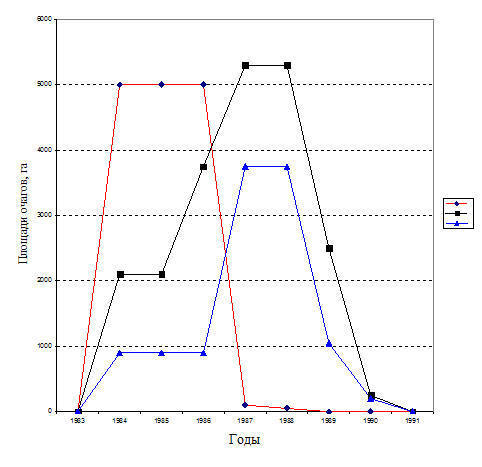
Рис.15. Особенности вспышки массового размножения шелкопряда-монашенки в лесах Свердловской обл. в 1982-1991 г.г.
♦ - Каменск-Уральский лесхоз, ■ – Камышловский лесхоз, ▲ - Сухоложский лесхоз
Новая вспышка массового размножения шелкопряда-монашенки в Свердловской области возникла в 1984-1990 гг.. В 1983 году мелкие, локальные очаги были обнаружены в сосновых лесах Каменск-Уральского, Камышловского и Сухоложского лесхоза на незначительной площади (рис. 15).
В этот период наблюдался интенсивный рост численности популяции в этих лесхозах. Поэтому на следующий 1984 г. площадь очагов значительно увеличилась. Наиболее крупные по площади очаги в этом году возникли в сосняках Каменск-Уральского лесхоза (рис. 15). Их общая площадь достигла 5000 га. В Камышловском лесхозе возник очаг на меньшей площади - 2100 га, а в Сухоложском лесхозе - 860 га (рис. 15).
Топографически все очаги, возникшие в 1984 году, локализованы в близлежащих районах в юго-восточной части Свердловской области, в северно-лесостепной (колочной) ново-березовой предлесостепной подзонах (Колесников, 1961, 1973). Очаги большей площадью проявляют явно выраженную тенденцию к локализации либо в подзоне северной лесостепи, либо непосредственно примыкая к ней (Колтунов, 1998).
Так же, как и очаги массового размножения шелкопряда-монашенки в Челябинской области очаги в период вспышки 1972-1980 гг. по продолжительности можно дифференцировать на два типа: длительно действующие и коротко действующие.
К первому типу относятся наиболее крупные по площади очаги в сосновых лесах Каменск-Уральского и Камышловского лесхозов. Продолжительность их существования составила 5-6 лет. Короткоживущие очаги выявлены нами в сосновых лесах Сысертского и Тугулымского лесхозов. Они возникли, соответственно, в 1986 и 1987 годах и продолжительность их существования составила лишь 3 года, площадь очагов в Сысертском лесхозе составила в 1987 году 2277 га, в Тугулымском - 3134 га. В 1986 и 1987 годах естественный ход динамики вспышек массового размножения шелкопряда-монашенки был нарушен обработками очагов с наиболее значительной опасностью полной дефолиации дендробациллином с добавлением дециса. В 1986 году обработки проводились на площади 1710 га, что не превышает 12,9% от общей площади очагов в этом году (13160 га). Тем не менее, даже без обработки эти очаги затухли бы в 1990 году, когда наблюдалось массовое затухание вспышки в Челябинской области (рис. 9-11). Поэтому, имеются достаточные основания отнести последнюю группу к категории коротко действующих.
Как следует из приведенных выше данных, наиболее крупные по площади очаги формировались, преимущественно, в южной подзоне лесостепи и в северной части степной зоны Челябинской области. В горно-лесной зоне Челябинской и Свердловской областей очаги образуются очень редко, и составляют небольшую площадь. Как правило, средняя степень дефолиации крон сосны в этих очагах также менее значительна, чем в очагах других природных зон с более благоприятными климатическими условиями и наиболее интенсивным характером протекания засух.
Сравнительный анализ очагов в Челябинской и Свердловской области показывает, что общая площадь очагов в Челябинской области была в 2,4-2,6 раз выше, чем площадь очагов в Свердловской области. Это свидетельствует о том, что на северной границе ареала зауральской популяции шелкопряда-монашенки в, среднем, климатические условия менее благоприятны, а уровень энтоморезистентности сосны обыкновенной выше, чем в степной и степной зонах.
Как показали результаты, длительно действующие очаги массового размножения шелкопряда-монашенки в сосняках Зауралья характеризуются незначительной площадью и слабой дефолиацией крон. Общая площадь этих очагов не превышает 10% от площади сосновых лесов вне очагов (Колтунов, 1996). Пространственно они локализуются в разных районах Челябинской области и не имеют строгой приуроченности к какому-либо тому, или другому типу лесорастительных условий. Чаще они приурочены к границам ареала. Они возникают в древостоях сосны с относительно высокой энтоморезистентностью.
Следует отметить, что Еткульский район Челябинской обл., который постоянно является местом локализации длительно действующих мелких очагов шелкопряда-монашенки одновременно служит и относительно оптимальной зоной для формирования очагов массового размножения зауральской популяции непарного шелкопряда (Колтунов, 1996, Колтунов и др., 1998). Последний, как отмечалось выше также не образует в березовых лесах Еткульского района крупных очагов в течение четырех вспышек массового размножения (Колтунов, 1993; Колтунов и др., 1998).
Детальное изучение особенностей топографии массивов сосновых лесов в лесхозах с минимальными площадями очагов шелкопряда-монашенки показало, что сосновые насаждения располагаются часто в виде изолированных массивов. В районах с особенно крупными по площади очагами массового размножения этого фитофага сосновые леса располагаются более крупными массивами, на близком расстоянии.
По нашему мнению это может быть важным фактором, влияющим на уровень реализации потенциала вспышки, так как в крупных лесных масссивах складываются более благоприятные условия для заселения новых участков путем миграций. Следовательно, особенности пространственой структуры сосновых лесов оказывают существенное влияние на уровень реализации потенциала вспышки у шелкопряда-монашенки. Поэтому сильная пространственная разобщенность сосновых боров на юге Челябинской области в зоне степи ограничивает потенциал реализации вспышки и площадь очагов.
Как показали результаты анализа изменения годичного радиального прироста сосны в очагах (Колтунов, 1998) именно реакция древостоев на фактор абиотического стресса (весенне-летние засухи) является ключевым фактором, инициирующим подъем, численности и возникновение вспышки у шелкопряда-монашенки в Зауралье. Следовательно, и шелкопряд-монашенка и непарный шелкопряд имеют одинаковый механизм возникновения вспышек.
Сравнительный анализ площадей очагов массового размножения этого фитофага в Свердловской области с 1950 по 1995 г.г. показал, что интенсивность вспышек резко отличалась между собой (рис. 14). Наиболее интенсивная вспышка массового размножения и самые крупные по площади очаги с сильной дефолиацией наблюдались в период вспышки 1952- 1956 г.г. (рис. 13, 14). Общая площадь очагов в период эруптивной фазы составляла, в среднем, 95.178 га сосновых лесов. Второй по интенсивности была вспышка массового размножения шелкопряда-монашенки в 1983-1991 г.г.. Ее площадь составляла 17.000 га. Менее интенсивной была последняя вспышка массового размножения (2003-2009 г.г.). Ее максимальная площадь не превышала 8200 га. Наименее крупной вспышкой была вспышка массового размножения 1973-1980 г..г. Общая площадь очагов в период максимума вспышки составляла - 5424 га сосновых лесов (рис. 13).
Таким образом, для наиболее интенсивных вспышек массового размножения зауральской популяции шелкопряда-монашенки на северной границе очагов массового размножения этого фитофага характерна минимальная продолжительность, очень интенсивный рост плотности популяции и площади очагов. По этому параметру данный тип вспышек целесообразно отнести к быстротекущему типу. В то же время длительно текущие вспышки массового размножения шелкопряда-монашенки на северной границе очагов массового размножения этого фитофага характеризовались менее интенсивным характером подъема численности в целом были либо незначительными, либо средними по площади очагов.
Интересно отметить, что последняя вспышка массового размножения зауральской популяции сибирского шелкопряда на северной границе очагов этого фитофага, которая была катастрофичной по площади погибших лесов (более 26.000 га темнохвойных лесов) также относилась к типу быстротекущих вспышек (Колтунов и др., 1998).
Учитывая сходство факторов возникновения очагов у сибирского шелкопряда, непарного шелкопряда и шелкопряда-монашенки целесообразно также было провести сравнительный анализ интенсивности вспышек массового размножения этих фитофагов за последние 45 лет.
Результаты сравнительного анализа убедительно показали, что наблюдается практически полное совпадение вспышек по их интенсивности, общей площади очагов у всех трех видов насекомых-фитофагов. Наиболее крупные по площади очагов вспышки наблюдались у всех этих видов насекомых-фитофагов в 50 годы, второй по площади очагов была также вспышка и наименее крупной - вспышка в 70 -е годы.У сибирского шелкопряда в Зауралье в 70 -е годы вспышки вообще не отмечалось (Колтунов и др., 1998).
Несомненно, полученные результаты однозначно свидетельствуют о том, что интенсивность вспышек и общая площадь очагов у всех трех видов насекомых-фитофагов, принадлежащих к одной группе по типу популяционной динамики (эруптивному), детерминируется общими для всех популяций факторами. Этот фактор: различная интенсивность засух. Очевидно, что фактор - цикличность наступления особенно сильных засух, имеет 3 летний период. Чередование "слабых" и "сильных" засух отмечались и другими авторами (Ильинский, 1958; Рожков, 1965). Но для некоторых видов насекомых-фитофагов этого ранее не отмечалось.
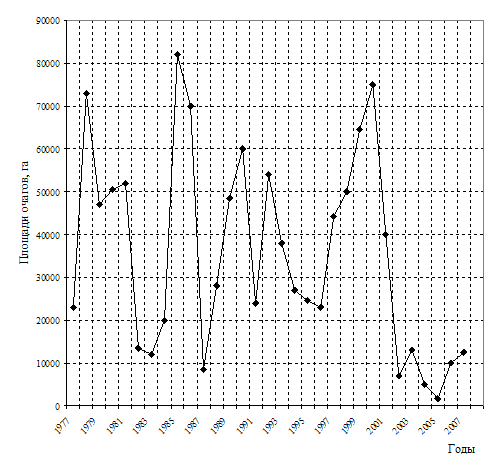
Рис. 16. Динамика площади очагов массового размножения шелкопряда-монашенки в лесах Российской Федерации (по данным Рослесхоза, 2007).
Как показали результаты, полученные нами ранее, для успешной реализации вспышки необходима синхронизация засух с периодом, предшествующим подъему численности популяции, продромальной и эруптивной фазам, а, также, определенный тип весенне-летних засух, их характер, интенсивность и продолжительность.
Это обусловлено тем, что, несмотря на наличие высокого биотического (вспышечного) потенциала у этой группы эруптивных видов лесных насекомых-филлофагов, уровень этого биотического потенциала не является постоянным, а в разные периоды популяционной динамики колеблется в очень широком диапазоне (Бахвалов, 2001; Колтунов, 2006). В, частности, в межвспышечный период он очень низок, в период вспышки очень высок.
Основным механизмом быстрого роста биотического (вспышечного) потенциала является способность группы эруптивных видов к быстрой адаптации популяции к изменению биохимического состава кормового субстрата. Именно различия в уровне и скорости адаптации детерминируют основные различия между эруптивными видами, способными к панзональным вспышкам и неэруптивными видами насекомых.
При этом адаптация может происходить как к кормовому субстрату, в насаждениях, подвергшихся воздействию фактора абиотического стресса (сильных весенне-летних засух), так и в направлении кормового субстрата после резкого изменения засушливых условий в экотопах на влажные, вследствие завершения засушливого периода.
Но, ранее нами было установлено, что временное снижение уровня энтоморезистентности в ответ на воздействие фактора абиотического стресса на насаждения происходит не у всей ценопопуляции, а только у той ее части, которая проявила наиболее сильный отклик на воздействие фактора абиотического стресса (засуху).
И это, по нашему мнению, является одним из ключевых звеньев реализации механизма вспышек массового размножения и эруптивных видов лесных насекомых-филлофагов.
Только сильная реакция на фактор абиотического стресса у большей части насаждений и быстрая временная потеря энтоморезистентности дает возможность быстрого роста заселенности насаждений популяцией насекомых, приводит к значительному росту их выживаемости. Слабая реакция на фактор абиотического стресса, свойственная древостоям с высокой энтоморезистентностью (например, растущим в более влажных лесорастительных условиях) не сопровождается столь резкими изменениями биохимического состава кормового субстрата (хвои и листьев) и ростом выживаемости популяции насекомых, вследствие высокой смертности популяции при питании таким кормовым субстратом. При этом, после изменения условий (прекращения засух и воздействия фактора абиоитического стресса на насаждения) и резкой смены условий на влажные уровень энтоморезистентности древостоев, резко отреагировавших на абиотический стресс, вновь восстанавливается.
Поэтому способность эруптивных видов к быстрой адаптации к измененному биохимическому составу кормового субстрата в результате воздействия на насаждения фактора абиотического стресса (засух) и сильной реакции на этот фактор, инициирует параллельно, с ростом уровня адаптации и быстрый рост биотического (вспышечного) потенциала.
Механизм затухания вспышек обусловлен феноменом кризиса адаптации популяции к кормовому субстрату, сформировавшемуся в результате воздействия фактора абиотического стресса (засух) на чувствительные к этому фактору насаждения, отреагировавшие на этот стресс резким изменением биохимического состава корма в сторону улучшения. Кризис адаптации возникает вследствии необходимости адаптации популяции фитофага к биохимическому составу кормового субстрата, но невозможности мгновенной адаптации к нему. Это сопровождается резким возрастанием смертности популяции, снижением выживаемости и резким уменьшением заселенности насаждений популяцией насекомых-филлофагов. Дальнейшие последствия адаптации популяции к изменению качества кормового субстрата тесно зависят от особенностей рельефа и параметров эдафотопа. В пессимальных условиях (пониженные элементы рельефа и увлажненные условия) уровень биотического потенциала популяции шелкопряда-монашенки остается низким.
Именно поэтому искусственное заселение таких насаждений яйцекладками никогда не сопровождается заметным ростом экологической плотности популяции фитофага. И лишь в оптимальных экотопах (на повышенных элементах рельефа) при локальном снижении влажности почвы и повышении температуры возможен незначительный локальный рост заселенности древостоев, который затем, при ухудшении экологических условий быстро прерывается.
Следовательно, детальный анализ особенностей популяционной динамики шелкопряда-монашенки в Зауралье за последние 50 лет и гидротермических условий в разные фазы популяционного цикла убедительно показывает, что механизмы и факторы возникновения вспышек массового размножения у шелкопряда-монашенки, сибирского шелкопряда и непарного шелкопряда совершенно идентичны.
Чаще всего очаги действуют в лесах Челябинской, Свердловской, Курганской,Тюменской, Новосибирской областей и Алтайского края. Во время последней крупной вспышки в 2000 году очаги были отмечены на общей площади 73,7 тыс. га в насаждениях 7-ми субъектов Российской Федерации, большая часть из них в Ульяновской (27,0 тыс. га) и Курганской (22,1 тыс. га) областях. Начиная с 2001 года, площадь очагов монашенки держится на уровне значительно ниже среднемноголетнего показателя (рис. 16).
Рекомендации по защите леса от шелкопряда-монашенки
В целом, многолетние исследования убедительно показали, что в благоприятных лесорастительных условиях сосна обыкновенная имеет достаточно высокий уровень энтомотолерантности. Именно по этой причине мы не рекомендуем планировать мероприятия по обработке очагов, если прогнозируется слабая и средняя дефолиация крон (до 50%). По нашему мнению, лишь точное прогнозирование ожидаемой 100%-ной дефолиации крон может быть достаточным обоснованием необходимости проведения мероприятий по обработке очагов.